This is the Part 2 of a three-part series of blog entries on the epigenetic’s of cancer and aging and how those two deadly dragons can be seriously slowed down or stopped with the assistance of plant polyphenols. The Part 1 blog entrytells the central story. It 1. identified similarities in the biological processes and epigenetic’s of cancer and aging, 2. identified therefore how common strategies might be found that address both cancer and aging. 3. described the process of Xenohormesis whereby stress response chemicals developed over millions of years in plants that respond to stresses and keep plants healthy can do the same in humans. 4. provided molecular explanations for the “causality” of cancer and aging, 5. described the processes in cancer and aging of epigenetic silencing of “good” genes and epigenetic activation of “bad” genes, 6. identified a 3 tiered “Pyramid” approach for chemoprevention of aging and cancer, 7. Iidentified the exact interventions involved in each layer of the Pyramid, and 8. Identified how the interventions in the three layers of the Pyramid can be integrated together. This Part 2 blog entry is concerned in more detail withthe silencing of good genes in aging and cancer and how polyphenols can prevent that. It lists naturally occurring phytochemicals that inhibit pathways that are known to be unregulated or dysregulated in cancer. The Part 3 blog entry in this Two-Dragon series is concerned: a. the “unsilencing of bad genes” with sirtuin decline and the harmful results in aging and cancer, and B. providing a master list of drugs and natural compounds for cancer chemoprevention. The Part 2 and Part 3 blog provide a series of appendices to the Part 1 entry, They’re published separately because of blog length considerations and because they are of interest in their own right.
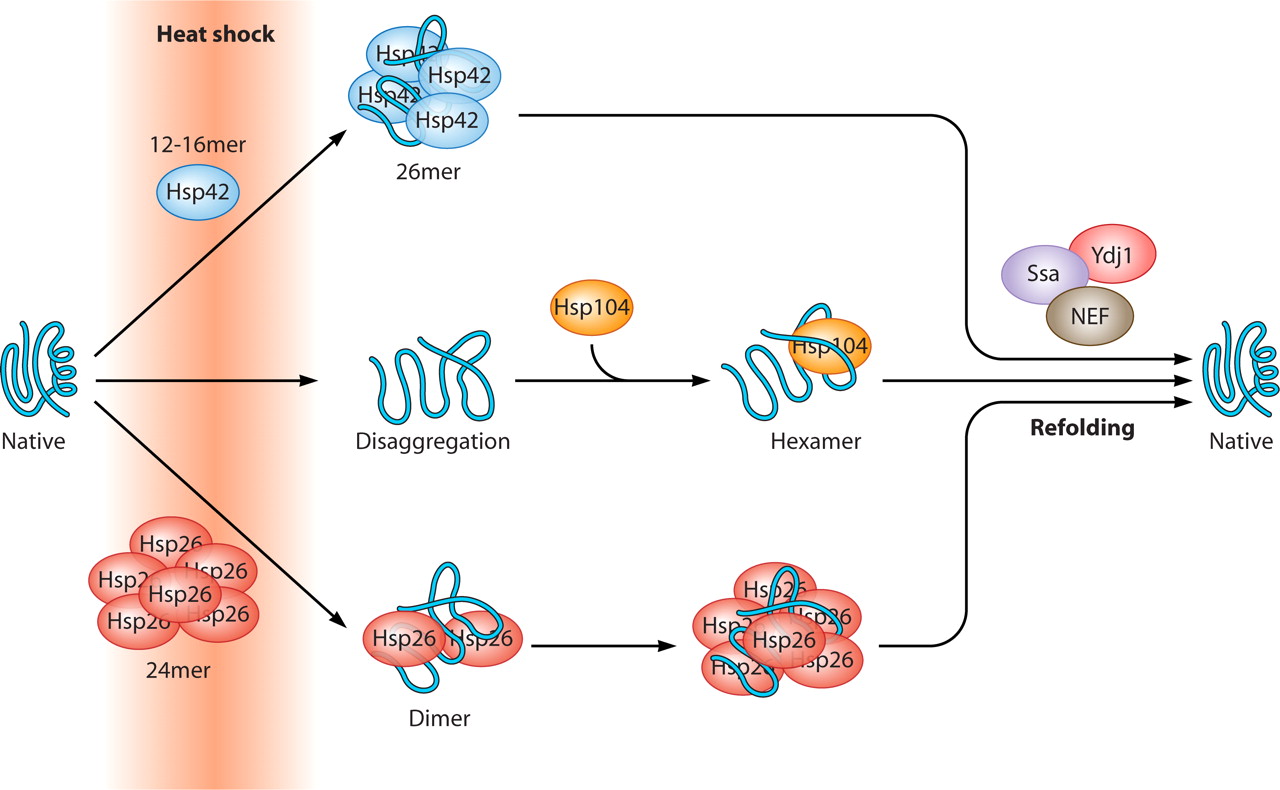
Part of Aging and Carcinogenesis is due to gene silencing: histone deacetylation, DNA methylation, & chromatin structure Δ: euchromatin è heterochromatin
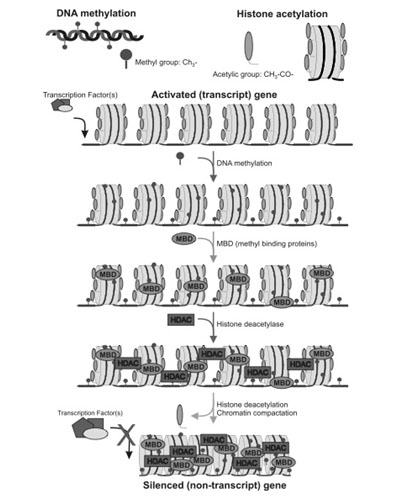
We now know that in many cases, gene inactivation is NOT due to a bona fide DNA mutation. Instead, the non-mutated gene can be silenced by an epigenetic mechanism that involves histone deacetylation of the histone proteins surrounding active genes (euchromatin), as well as DNA methylation of a cytosine-rich region of promoter sites (“start location”) on the gene. These cytosine-rich areas are called CpG islands and the cytosine bases can be methylated by the enzyme DNMT. Thousands of “good genes” must remain in the euchromatin state to allow for gene expression to occur. When histones are acetylated, transcription factors can reach the DNA because of the “open structure” of the histones. When the promoter region has a hypomethylated CpG island, the transcription factor can bind to the DNA promoter region. Transcription factors migrate into the nucleus in response to various cellular signaling pathways. are “silenced”, they cannot be turned on by cellular signals, even if the gene is not mutated.
The following discussion explains how gene silencing occurs and then gives some specific examples of gene silencing in aging and cancer. Further, it goes on to illustrate how specific plant polyphenols can be used to unsilence anti-aging and anti-cancer genes and to silence pro-cancer and pro-aging genes.
Gene silencing by epigenetic mechanisms is an ordered series of events that normally starts with the methylation of cytosine residues. Scientists studying gene silencing have observed two distinct kinds of CpG methylation: one that is age-related that occurs in all tissues as a function of aging, and a cancer-related pattern of methylation that only occurs in cancerous cells. It is important to point out that with aging, there is a global loss of methylation at sites other than CpG islands. The hypermethylation of CpG islands is the exception to this universal decline in DNA methylation with aging. This can be explained in part by the fact that there are different DNA methyltransferases for these different regions of the genome. DNMT1 is responsible for the maintenance of hemimethylated DNA, whereas DNMT3a and 3b are responsible for the methylation of unmethylated DNA. For this reason, it appears that DNMT3a and 3b are responsible for gene silencing.
Methyl-binding proteins and Histone deacetylases (HDACs)
After the CpG residues have been methylated by DNMT, methyl binding proteins attach to the methyl groups, creating the first layer of protein-based silencers. Methyl binding proteins then recruit histone deacetylases (HDACs) to the histone proteins.
Histone deacetylation and chromatin compaction
Once the HDACs have been recruited, they remove specific acetyl groups from the lysine side chains on the 9th amino acid of the Histone 3 protein. (H3K9 deacetylation). Once the histone groups have been removed, the nucleosomes can be compacted into a tight configuration called heterochromatin.At this point, the gene is effectively silenced and reactivation of the gene is difficult.
Preventing Histone-based Gene Silencing : Inhibiting histone deacetylation of Lysine 9 on Histone 3 (H3K9) with polyphenols
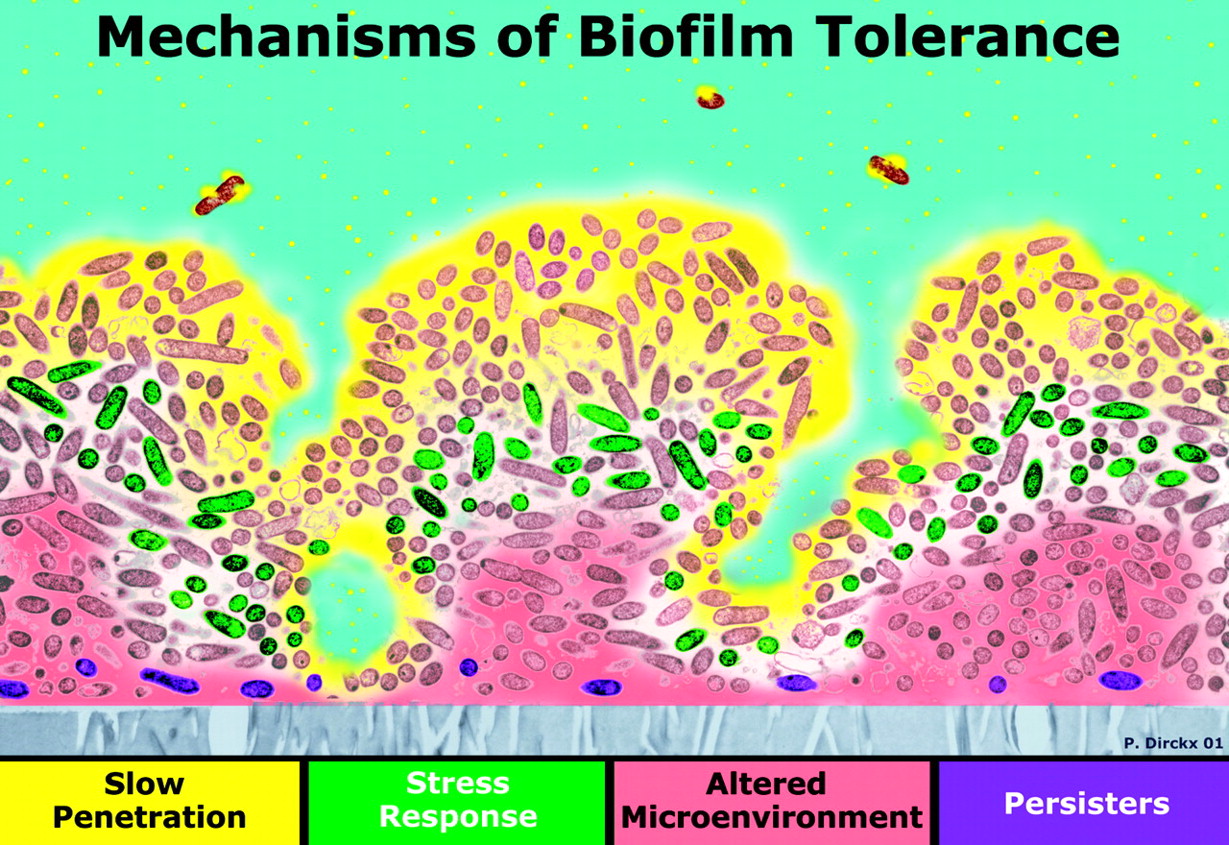
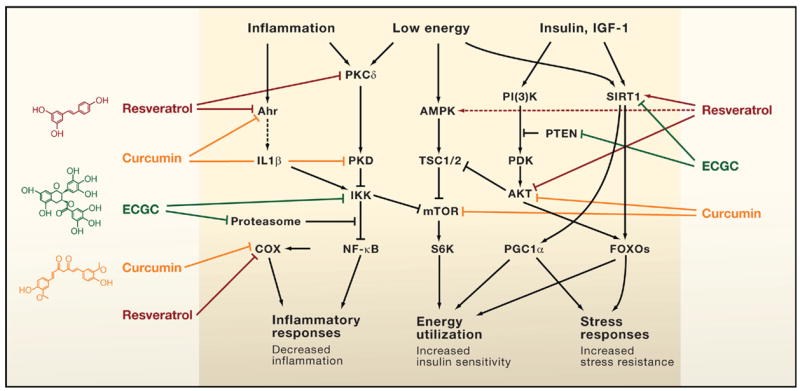
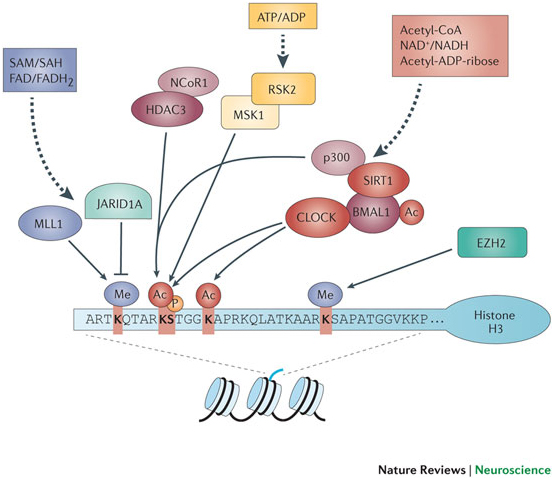
Aging and cancer exhibit epigenetic silencing of genes that should NOT be silenced. Since only 50% of genes have CpG islands at their promoter sites, DNA methylation is not as common a method of gene silencing as histone-based gene silencing, which can occur for almost all genes. Histone-based gene silencing is best understood as an imbalance in the equilibrium between histone acetylation (by HAT) and histone deacetylation (by HDACS). The diagram below illustrates this and how plant polyphenols can inhibit the equilibrium both ways, since some of the polyphenols are both HDAC inhibitors and HAT inhibitors. HDAC inhibition is a way of preventing gene silencing of euchromatin, whereas HAT inhibition is primarily a way of preventing the activation (expression) of repetitive DNA, which normally is silent. The transcriptional activation of repetitive DNA is increasingly being studied since it seems to be the consequence of the global decline in DNA methylation that occurs with aging.
The figure above shows the representation of histone modifications (acetylation and deacetylation) by the phytochemicals derived from different food source. Phytochemicals like EGCG, genistein and curcumin play important role in inhibition of histone acetylation by inactivating histone acetyl transferase enzyme. Some other phytochemicals like sulforaphane, curcumin, genistein, phenyl isothiocynate, organosulfur compound, resveratrol and indol-3-carbinol inhibits the deacetylation of relaxed chromatine by inactivating histone deacetylase enzyme. Here is a list of several other phytochemicals that alter histone modifications which are not covered in this figure:
Compound
Source
Function
Anastatins
Egyptian medicinal herb
(-) HDAC
Apicidin
Fungal metabolite
(-) HDAC
n-butyrate
metabolite from gut bacteria fementing fiber
(-) HDAC
Caffeic acid
green coffee beans
(-) HDAC
Chlorogenic acid
green coffee beans
(-) HDAC
Cinnamic acid
cinnamon
(-) HDAC
Hydroxycinnamic acid
cinnamon
(-) HDAC
Equol
created when gut bacteria metabolize diazen, an ingredient in soybeans
(-) HDAC
Erucin
broccoli, brussel sprouts
(-) HDAC
Phenolic acids
dried fruits
(-) HDAC
Plumbagin
fly venus trap extract
(-) HDAC
Selenium
brazil nuts, meat
(-) HDAC
Trapoxin
fungus Helicoma ambiens
(-) HDAC
Zerumbone
ginger extract
(-) HDAC
Preventing DNA methylation-based Gene Silencing: Inhibiting DNA methylation at the CpG islands found in promoter sites with polyphenols
50% of the promoter sites in human genes have a region that is rich in cytosine bases. This region is called a “CpG island” and is important in the transcriptional activation of gene expression by transcription factors. In genes that have a CpG island (50% of the total), gene silencing can occur by methylation of the cytosine bases in the CpG island. This prevents gene transcription. The enzyme responsible for methylating the cytosines is called DNA methyltransferas. This is normally the first step in gene silencing and occurs before histone deacetylation. Many natural products including green tea, soybeans, garlic, tomatoes, watercress, curry powder, broccoli, brussel sprouts, cabbage, red grapes, and olives prevent this step in the silencing of “good genes”. The diagram below illustrates this.
DNA methylation is a biochemical process that is essential for the development of higher organism. Some dietary phytochemicals are reported to inhibit the methylation of deoxycytidine at DNA promoter sites. Hypermethylation of cytidine by DNMTs usually results in transcriptional gene silencing and gene inactivation. Several phytochemicals derived from different food source such as: resveratrol from graps and berries, curcumin from turmeric tea phenols from tea leaves, genistein from soybeans, sulforaphane from broccoli, phenethyl isothiocynate from cauliflower, organosulfur compounds from garlic, quercetin from citrus fruits and lycopene from tomato act as dietary inhibitors of DNA methyl transferases and also alter gene expression via epigenetic mechanisms. Here are some other natural DNMT inhibitors:
Compound Source Effect
Caffeic acid
Green coffee beans
(-) DNMT
Chlorogenic acid
Green coffee beans
(-) DNMT
Garcinol
Mangostein rinds
(-) DNMT
Hesperitin
Citris rinds
(-) DNMT
Hydroxycinnamic a.
Cinnamon
(-) DNMT
Kazinol Q
Formosan plants
(-) DNMT
Luteolin
Tomato skins
(-) DNMT
Lycopene
Tomato skins
(-) DNMT
Myricetin
Grapes, berries, veges
(-) DNMT
Naringenin
Grapefruits, oranges, & tomato skins
(-) DNMT
Phloretin
Apple skins
(-) DNMT
Quercetin
Apple skins, onions
(-) DNMT
Selenium
Brazil nuts, meats
(-) DNMT
Other methods of Gene silencing – Histone protein trimethylation and Polycomb protein co-repressor silencing of genes
There are other mechanisms of epigenetic silencing not covered in this appendix, but should be mentioned for the sake of completeness. When histone protein tails are deacetylated, the lysine side chains are typically methylated. Single methyl groups or two methyl groups are associated with down regulation of gene expression but not silencing. Trimethylation of the histone protein lysine side chains is typically associated with gene silencing. Polycomb proteins are another form of gene silencing that has been evolutionarily conserved from lower organisms that do not methylate DNA. The significance of Polycomb protein-based gene silencing is not as clear has the other epigenetic mechanisms described in this Two-Dragon series of entries. Nevetheless, this Polycomb silencing is important to understand and occasionally comes up in discussions of gene silencing.
Gene Silencing by non-coding RNA – miRNA silencing of tumor suppressor genes and activation of oncogene expression
A third method by which genes can be silences is by the inhibition of messenger RNA (mRNA) by non-coding RNA called miRNA (also called microRNA). These miRNA bind to mRNA, making them incapable of creating a protein, or increasing the degradation of the mRNA, thereby eliminating the possibility that the mRNA can create a protein. This means that the gene may be transcribed (i.e. a messenger RNA is made), but the mRNA transcript is not “read” and translated into a protein in the ribosomes. This method of gene silencing can also be used to activate or increase the expression of an oncogene. Most oncogenes are actually good and necessary genes expressed with normal cellular function, but become overexpressed with cancer. When this occurs, such a gene is referred to as an “oncogene”. The diagram below illustrates how miRNA can inhibit a tumor suppressor mRNA and activate an oncogene. Several naturally occuring foods have active ingredients that prevent tumor suppressor mRNAs from being silenced by miRNA, as well as preventing oncogene mRNAs from being translated into proteins. These natural foods include berries, green tea, soybeans, broccoli, and curry spice.
How to Suppress Oncogenes and Activate Tumor Suppressor Genes
with Tea, Tofu, Food, and
Dietrary polyphenols can affect microRNA (miRNA) expression. miRNAs are noncoding RNAs that regulate gene expression after the gene is transcribed into a messenger RNA (mRNA). miRNA are transcribed in the nucleus into primary miRNA (pri-miRNA) which is further cleaved by Drosha into precursor miRNA (pre-miRNA). Pre-miRNA is exported from nucleus to the cytoplasm and further processed by Dicer into miRNA duplex. Single strand of miRNA duplex (also called mature miRNA) leads this complex to mRNA cleavage or translation repression, which is dependent on miRNA:mRNA complementarity. Dependent on various factors, miRNA can have either an oncogenic role (called onco-miRNAs) if the target mRNA is a tumor suppressor gene, or a tumor suppressive role (tumor-suppressor miRNAs) if the target molecule is an oncogene. Dietary polyphenols can impact expression level of miRNAs and participate in gene expression regulation.
How to “Reactivate” your Silenced Genes
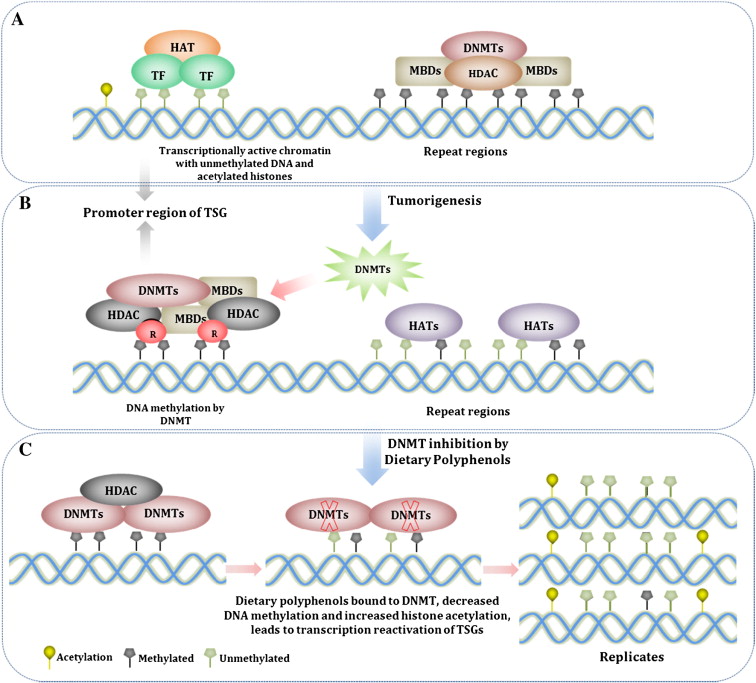
The epigenetic silencing of “good genes” is not irreversible. Natural compounds, when used on a regular daily basis can reverse this phenomena and reactivate silenced tumor suppressor genes. The diagram below illustrates this:
A. Inhibit HDACs – Histone deacetylases are enzymes found in the cell nucleus that remove acetyl groups from the histone proteins. This inactivates the transcriptional machinery. This is the most common way that dietary polyphenols turn genes back on. This step usually precedes DNMT activity.
B. Inhibit HAT – Histone acetyltransferases are enzymes that reacetylate histone proteins, opening the chromatin structure of DNA, allowing the genes to be reached by transcription factors. At first glance, you would think that HAT activation would be good, rather than inhibition. Unfortunately, HATs also have a “dark side” to their function, especially with regards to repetitive DNA sequences. Normally, repetitive DNA is silenced and should NOT be transcribed, since this results in genomic instability. Unfortunatley with aging and with cancer, there is a global decline in DNA methylation (opposite of promoter regions where there is an increase in DNA methylation). When HATs bind to demethylated regions of repetitive DNA, they acetylate the repetitive DNA, allowing transcription of the repeat regions. This can contribute to tumorigenesis and genomic instability. There are several natural compounds have been shown inhibit HATs and thereby prevent the “unsilencing” of repetitive DNA. This includes allspice (eugenol, ericifolin), cashew nut oil (anacardic acid), green tea (EGCG) soybeans (genistein), and turmeric (curcumin).
C. Inhibit DNMTs - DNMT activity is blocked by dietary polyphenols by forming hydrogen bonds with amino acids (Pro, Glu, Cys, Ser, and Arg) in the catalytic pocket of DNMT. Newly synthesized DNA strands are semi-methylated after the first round of DNA replication and become progressively more demethylated after several rounds of replication due to the dilution effect. This is why life long dietary intake of polyphenols that inhibit DNMT is so important.
Specific Examples of Genes known to undergo epigenetic silencing
It is important to point out that not all genes have CpG islands. In humans, approximately half of the all genes have CpG islands, but the very important genes involving “housekeeping genes” and “tissue-specific genes” seem to have a disproportionate share of CpG islands. These important genes are silenced by a combination of histone deacetylation and CpG island DNA methylation at the promoter sites. This results in the inability of a transcription factor to access the gene and to bind to the promoter site. With aging, many genes are silenced this way without any mutation of the DNA. There are some specific genes that are silenced via this epigenetic mechanism that play a huge role in aging. Here are some of them:
1. DNA repair gene inactivation by epigenetic silencing – Four examples of this are the genes hMLH1, MGMT, WRN, and BRCA1
hMLH1 – this an important gene for DNA mismatch repair, which is the way that microsatellite-unstable regions are repaired to avoid them becoming bigger. Gene inactivation by CpG island hypermethylation has been observed at the promoter site for this gene
MGMT – this is an important gene for fixing mutant guanine bases that become chemically modified by a methyl or alkyl group. This modification results in the “G” being read as a “A” (i.e. creating a G-to-A single nucleotide polymorphism). MGMT removes the pro-mutagenic guanine residue. Unfortunately, this gene is inactivated by CpG island hypermetylation with aging.
WRN – this is the gene responsible for Werner’s syndrome. This gene codes for the WRN protein which has helicase and exonuclease activity. With aging, the CpG island at the promoter site for this gene is hypermethylated, effectively silencing this gene. As a consequence, the manifestations are a progeroid phenotype and increased risk of cancer due to extreme sensitivity to DNA damaging drugs or toxins.
BRCA1 – this gene is the most common gene that is mutated with hereditary breast cancer. However in sporadic cases of breast and ovarian cancer, the BRCA1 gene can be silenced by CpG island hypermethylation in the promoter site of this gene.
2. Progeroid syndromes and atypical Werner’s syndrome– inactivation by epigenetic silencing. Two examples of accelerated aging genes – Lamin A/C and WRN
Laminin A/C – Mutations in the Laminin A/C gene produce a syndrome called Hutchinson’s Gilford progeria. Although most progeroid syndromes are due to DNA mutations, we now know that a progeroid-like phenomena can be acquired due to epigenetic silencing of the same gene that is mutated in the hereditary form of HG progeria. The Laminin A/C gene codes for two different laminin proteins that make up the scaffolding just inside the nuclear double membrane. When this gene is hypermethylated at the promoter site, a syndrome called “atypical Werner’s syndrome” occurs.
WRN – this is called the “Werner gene” mentioned above. It is mutated in classic Werner’s syndrome (WS). Patients with WS develop accelerated aging and manifest cataracts, type II diabetes, osteoporosis, arteriosclerosis, and cancer at an earlier age and at increased incidence. Cases of WS where the only problem is CpG island hypermethylation at the promoter site site. Again, this is where we need a “DNA methylome” for proper diagnosis.
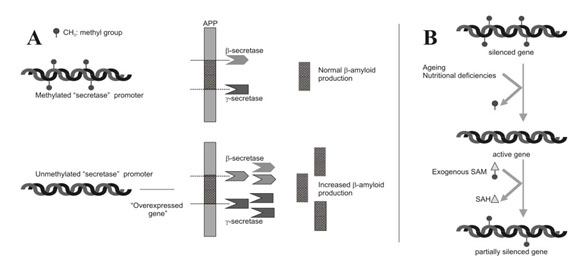
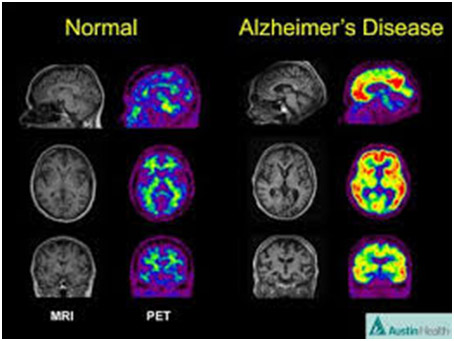
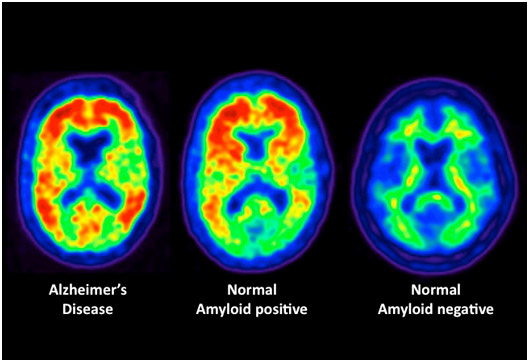
3. Alzheimer’s disease – Epigenetic “unsilencing” of PS1 and BACE genes. Alzheimer’s disease appears to be a multifactorial problem that is age-related. Epigenetic silencing is only one of the explanations for the disease. Several explanations that involve epigenetic mechanisms may be involvedwith AD – homocystein/SAM dysregulation, DNA methylation/demethylation of genes, and the effects of folate and B12 deficiencies on the methylation status of genes involving AD.
Two Epigenetic Explanations for Alzheimer’s Several studies have shown that both the gamma-secretase gene and the beta-secretase genes are under the control of DNA methylation of their promoter sites. Beta-secretase cleaves APP in the wrong place, creating amyloid-beta. DNA demethylation causes the overexpression of PS1 and BACE genes, which result in an increase in amyloid-beta.
Nutritional deficiencies can also explain AD. In vitro studies of neuroblastoma cells show that nerve cells cultured with a B12 or folate deficiency result in the over production of amyloid-beta. Adding B12 or folate can reverse the increase in amyloid-beta. Exogenous SAM can also restore the methylation pattern and silence the gene in a physiologic way.
Naturally occurring phytochemicals that inhibit pathways that are known to be unregulated or dysregulated in cancer.
Big Pharma is developing drugs to block most all of these pathways. You can block them yourself by going for a walk in the jungle, the forests, or the mountains.
You may notice a recurrent theme in the following table: the best health-producing natural plant substances may be found in the dumpsters of industrial plants that produce commodity food and drink substances like orange juice, wines, tomato ketchup, olive oil and applesauce. This is because the most-powerful naturally-produced stress resistant phytochemicals substances occur in the skins and rinds that are normally removed in the course of making commodity products. Skins are where oranges, grapes, tomatoes, olives, grapefruit, nuts, apples, and many other plants foods have found an evolutionary need to concentrate their chemical stress defenses. If we want to take advantage of the xenohormetic properties of these substances we have to find them where they are – today often in the dumpster.
Pathway Inhibited (or activated) – CompoundNatural Sources of Compound
Nonspecific Tyrosine Kinase Inhibitors
genistein — soybeans – maybe we can get these from soy milk manufacturing plants
resveratrol — polygonum cuspidatum roots, Japanese knotweed, red grape skins
quercetin – apple skins, onions, etc. – maybe we can get these from apple juice plant garbage
luteolin – tomato skins – maybe we can get these from Campbell’s tomato soup plant
The tables and compilations of data in this and the other blog entries in the Two Dragons series are intended to be illustrative of the main points to be made. They are compiled from various sources, in most cases are incomplete, and may contain occasional errors.
MEDICAL DISCLAIMER: FROM TIME TO TIME, THIS BLOG DISCUSSES DISEASE PROCESSES. THE INTENTION OF THOSE DISCUSSIONS IS TO CONVEY CURRENT RESEARCH FINDINGS AND OPINIONS, NOT TO GIVE MEDICAL ADVICE. THE INFORMATION IN POSTS IN THIS BLOG IS NOT A SUBSTITUTE FOR A LICENSED PHYSICIAN’S MEDICAL ADVICE. IF ANY ADVICE, OPINIONS, OR INSTRUCTIONS HEREIN CONFLICT WITH THAT OF A TREATING LICENSED PHYSICIAN, DEFER TO THE OPINION OF THE PHYSICIAN. THIS INFORMATION IS INTENDED FOR PEOPLE IN GOOD HEALTH. IT IS THE READER’S RESPONSIBILITY TO KNOW HIS OR HER MEDICAL HISTORY AND ENSURE THAT ACTIONS OR SUPPLEMENTS HE OR SHE TAKES DO NOT CREATE AN ADVERSE REACTION
This is the Part 3 of a three-part series of blog entries on the epigenetic’s of cancer and aging and how those two deadly dragons can be seriously slowed down or stopped with the assistance of plant polyphenols. The Part 1 blog entrytells the central story. It 1. identified similarities in the biological processes and epigenetic’s of cancer and aging, 2. identified therefore how common strategies might be found that address both cancer and aging. 3. described the process of Xenohormesis whereby stress response chemicals developed over millions of years in plants that respond to stresses and keep plants healthy can do the same in humans. 4. provided molecular explanations for the “causality” of cancer and aging, 5. described the processes in cancer and aging of epigenetic silencing of “good” genes and epigenetic activation of “bad” genes, 6. identified a 3 tiered “Pyramid” approach for chemoprevention of aging and cancer, 7. Iidentified the exact interventions involved in each layer of the Pyramid, and 8. Identified how the interventions in the three layers of the Pyramid can be integrated together. The Part 2 blog entryis concerned in more detail withthe silencing of good genes in aging and cancer and how polyphenols can prevent that. It lists naturally occurring phytochemicals that inhibit pathways that are known to be unregulated or dysregulated in cancer. This Part 3 blog entryis concerned with: A) the silencing of bad genes and repetitive DNA to avert harmful consequences in aging and cancer, and B) providing a master list of drugs and natural compounds for cancer chemoprevention. The Part 2 and Part 3 blog entries provide a series of appendices to the Part 1 entry, They are published separately because of blog length considerations and because they are of interest in their own right.
PART A – Silencing repetitive DNA sequences
The Sound of Silence– How Leonard Guarante discovered Sir2, the repetitive rDNA silencer, in Baker’s yeast
A good way to present this is by taking a little romp through the history of actions of sirtuins.
Working with Baker’s yeast at MIT, back from 1997 to in 2004 Dr. Leonard Guarante screened yeast colonies for long-lived strains and discovered that a single mutation in the gene SIR4 caused the protein Sir2 to gather at the most repetitive DNA sequence in yeast – the rDNA repetitive gene sequence where there are about 100 copies of this gene for ribosomal proteins(ref)(ref). Unfortunately, this repetitive area is unstable and the gene copies tend to recombine with each other. This is called “genomic instability” and is seen with many other genes in humans (humans do not have an unstable rDNA area, however). Recombination of repetitive DNA is a major cause of human cancer and other human diseases such as Huntington’s disease(ref)(ref)(ref).
ERCs and gene silencing by Sir2– How an extra copy of SIR2 gene extended lifespan by 30% & 50%
In yeast, a very unusual event occurs when the rDNA copies combine with each other. With yeast aging, the mother yeast cell starts spinning off circular rings of this rDNA, which “pop out of the DNA”. These DNA rings are called “extrachromosomal rDNA circles” or ERCs(ref). When the mother yeast cell divides, these ERCs are also copied but all the ERC copies stay in the mother yeast cell. With successive cell divisions, the mother yeast cell becomes over-run with all these ERCs. These ERCs cause aging. The cellular energy and resources required to replicate all these ERCs eventually spells death for the mother yeast after about 20 cell divisions. When an extra copy of the SIR2 gene was added to the yeast cells, this extended the lifespan of the yeast by 30%. When the same experiment was done in nematodes, the extra copy of SIR2 extended lifespan by 50%. This was the first indication that SIR2 was a “longevity gene” and that the mechanism of life span extension was “gene silencing” of repetitive DNA(ref). So, in yeast at least something can be done about repetitive DNA and its life-shortening consequences, and that something involves SIR2. Of course, the question arose “What about mammals, including us?”
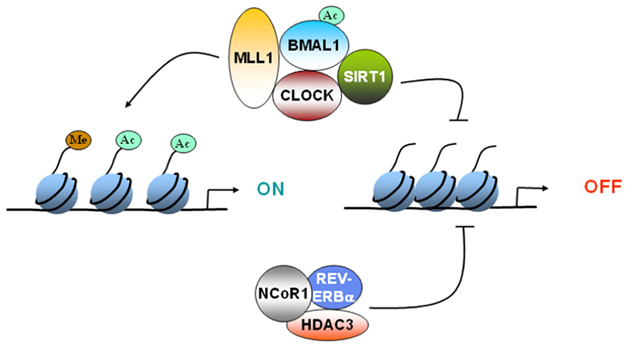
Histone deacetylation = Gene silencing
Once the Sir2 protein was discovered to be a gene silencer, the molecular mechanisms for this effect on gene expression was discovered to be the removal of acetyl groups from the histone proteins that shroud DNA. It was discovered that this Sir2 histone deacetylator could only do this if NAD+ was present. NAD+ was a cofactor required for decetylation to occur and high levels of NAD+ and low levels of NADH would stimulate Sir2. Since high levels of NAD+ reflected low energy levels in the cell, it was determined that these histone deacetylators were “energy sensors” responding to food (calorie) availability. This was the impetus to do caloric restriction studies in the Baker’s yeast.
Calorie Restriction ->Sir2 Activation ->Longevity
Caloric restriction(ref),(ref),(ref) had already been shown to extend lifespan in experiments done 70 years prior, but there were still major misconceptions about how caloric restriction could extend lifespan. The “conventional wisdom” at the time of Dr. Guarente’s yeast studies (2004) was that caloric restriction was due to slowing down metabolism, which would then reduce free radical production by mitochondria. (Lane, Ingram, Roth, Scientific American, 2004). Unfortunately, experimental studies did not support this “rate of metabolism theory”, since caloric restriction does not slow down metabolism in yeast, worms, and humans. This lack of experimental evidence for the “rate of metabolism theory” lead Guarante to consider Sir2 as a possible mediator of caloric restriction’s effects on longevity. With yeast caloric restriction activates Sir2 by two different mechanism:
1. PNC1 – PNC1 is an gene that codes for an enzyme that gets rid of nicotinamide, a molecule similar to vitamin B3. Nicotinamide inhibits Sir2, so getting rid of it activates Sir2
2. Respiration – mitochondrial energy production is called “respiration” and has a by-product called NAD+. Lowering NAD+ levels activates Sir2.
The final “proof” that Sir2 is required for the longevity effects of caloric restriction was done by “knocking out” the SIR2 gene, which eliminated the life span extending effects of caloric restriction in yeast. Adding an extra copy of SIR2 gene to fruit flies further extended the life span extending effects of caloric restriction(ref).
Resveratrol ->Sir2 Activation ->Longevity– the claims, the questions, and the vindication
When David Sinclair joined Dr. Guarente’s laboratory at MIT as a post doc, he did the now famous experiment that showed how resveratrol, a polyphenol found in grape skins and wine, activated Sir2 in yeast, which led to world wide attention to Sir2 as the ultimate “longevity gene” and red wine as the “fountain of youth”, explaining the French paradox (The paradox of why French people do not have as much heart disease as would be predicted, based on their lifestyle). Unfortunately, the results of this experiment could not be reproduced in other yeast laboratories and the scientific validity of this conclusion was questioned. Although few were questioning the longevity effect of resveratrol, some were questioning if this effect was mediated by Sir2. Over the past 7 years, this controversy has continued to smolder, with many claiming that the primary mechanism of action for resveratrol is AMPK activation. Earlier this year, however, David Sinclair and Leonard Guarente published a follow-up study that eliminated the questionable reagent used in the experiment. Without the questionable reagent, the results of the resveratrol experiment was the same as the original study. This paper has largely silenced the critics of the silencing ability of resveratrol on repetitive rDNA circle formation(ref).
There are activities, naturally occurring molecules and synthetic molecules that activate Sirtuins. These “sirtuin activating compounds” are called STACs. Here is a list of natural STACS and synthetic STACS. Please note that some activities or compounds only activate the enzyme, whereas other activities/compounds also activate gene expression of the SIRT enzyme:
Compound or Activity
Source/Mechanism of Action
Exercise (E)
E -> SIRT activity, E -> SIRT gene expression, but net effect is overall
Caloric Restriction (CR)
CR -> SIRT activity AND ->SIRT gene expression
Dietary polyphenols
polyphenols -> SIRT activity but do not -> SIRT gene expression
Resveratrol
4.6-5.2 fold -> in SIRT activity – not affected by lack of vitamin C
Fisetin
Quercetin
2.1-2.5 fold -> in SIRT activity – not affected by lack of vitamin C
ECg
1.85–1.91 fold -> in SIRT activity – not affected by lack of vitamin C
EC
0.99-1.09 fold -> in SIRT activity – not affected by lack of vitamin C
EGCg, EGC, myricetin, gallic acid
very weak SIRT activators that loose their effect without vitamin C
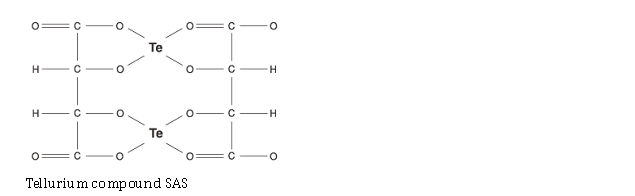
Tellurium compounds
-> SIRT activity AND -> SIRT gene expression
Synthetic analogs of resveratrol
synthetic STACS -> SIRT activity but do not -> SIRT gene expression
SRT 1720
100X more potent than resveratrol
SRT 2183
in development by Glaxo-Smith-Kline
SRT 1460
in development by Glaxo-Smith-Kline
Icarin
-> SIRT via MAPK kinase, does not -> SIRT gene expression
2-Deoxyglucose (2DG)
-> SIRT activity AND ->SIRT gene expression via -> association with redox s
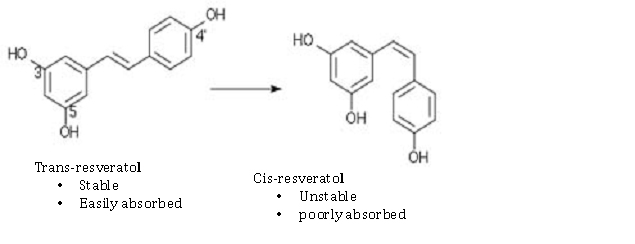
Here are some examples of the chemical configurations of these gene silencing compounds that work on Sirtuin activation or SIRT gene expression. As you can see, there are two enantiomers of resveratrol: trans-resveratrol and cis-resveratrol. The trans form is stable and easily absorbed. The cis form is unstable and poorly absorbed. For this reason, most dietary supplements are made with the trans form of the compound.
Differences in Caloric Restriction Mechanisms in Different Species
A very important distinction should be made in humans, where there are 7 different sirtuins. In humans, the sirtuin that is involved with metabolic control is SIRT1, which is the ortholog to the yeast SIR2. Because all of the 7 Sirtuins are “redox sensitive”, however, there will be other effects of caloric restriction besides the control of metabolic pathways. In other words, caloric restriction (CR) has traditionally been studied just with SIRT1 and the effects of SIRT1 on transcription factors. The other Sirtuins (SIRT2, SIRT3, SIRT4, SIRT5, SIRT6, and SIRT) are also “redox sensitive” but have completely different functions/effects. To date, SIRT1 has not been shown to have a longevity effect in humans. However recent studies have found that SIRT6 does have a longevity effect in humans. Another reason for the controversy about Sirtuins is that caloric restriction appears to have different pathways that mediate their effects in different organisms. As you can see below, mitochondria play a crucial role in CR. The following diagram illustrates some of these differences:
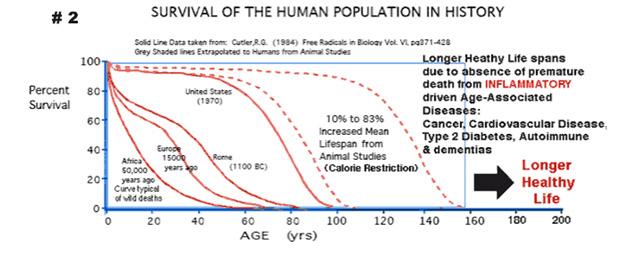
Calorie restriction increases life span in all model organisms, but has not shown a longevity effect in humans yet. Two longitudinal primate studies both showed positive effects on health, but only one of the two showed an increased life span effect. The diagram above from Guarante’s review article shows how it the yeast SIR2 gene othologs in mice and humans is the SIRT1 gene, but in nematodes sirtuins do not appear to be involved with the caloric restriction pathway. This may be due in part to the fact that repetitive DNA silencing is mostly due to polycomb protein group silencing mechanisms. In C. elegans, the SKN-1 gene mediates caloric restriction.
Mice
The pathway in mice shows that the increase in mitochondrial number and activity may work via mitochondrial sirtuins (SIRT3, 4, and 5) or by reducing reactive oxygen species (ROS). Resveratrol and CR may increase SIRT1 activity, which is part of an autoregulatory feedback loop that includes the enzyme endothelial nitric oxide synthase (eNOS). In this aspect, eNOS and SIRT1 have a “reciprocal causation” relationship, where SIRT1 deacetylates eNOS and eNOS activates SIRT1. In mice and probably humans, SIRT1 may increase PGC-1α activity by deacetyation of several lysine side chains, increasing the ability of PGC-1α to co-activate genes.
[Rodgers et al., 2005] and [Gerhart-Hines et al., 2007]). By this mechanism, SIRT1 can increase mitochondrial biogenesis. In humans, CR up regulates SIRT1, eNOS, TFAM, PGC-1 α, and AMPK. In humans, CR down regulates mTOR, the Insulin/IGF-1 pathway, and NF-kB. For these reasons, it is more difficult to give SIRT1 all of the credit for the effects of CR in humans.
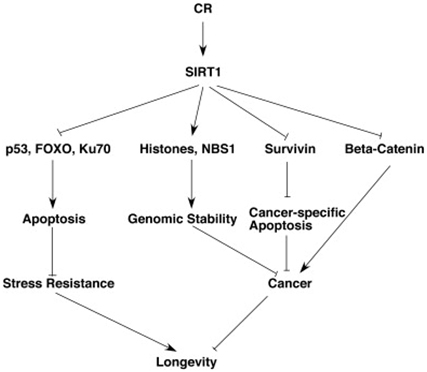
The Stress Resistance and Anti-cancer effects of Caloric Restriction and SIRT1
Although this blog is primarily about gene silencing of repetitive DNA, it is important to point out that caloric restriction is one of the most powerful cancer prevention strategies that has been found. CR is even more effective than polyphenol-based chemoprevention in in vitro cell studies and in vivo animal studies. This anti-cancer effect of SIRT1 is mediated via 5 mechanisms:
1. Survivin deacetylation: this induces cancer-specific apoptosis
2. β-catenin deacetylation: this prevents cancer growth
3. Notch deacetylation: this inhibits cancer angiogenesis
4. Histone & NBS1 deacetylation: this promotes genomic stability by gene silencing
5. Nrf2 and FOXO3 deacetylation: this increases gene expression of antioxidant enzymes
SIRT1 increases stress resistance by three separate mechanisms. SIRT1 deacetylates the FOXO transcription factors, thereby activating genes for anti-apoptosis mechanisms. SIRT1 also deacetylates p53, which makes it less likely to induce apoptosis. SIRT1 also activates Ku70, which activates DNA repair mechanisms. As a result of the 6 pathways mentioned above, SIRT1 increases longevity by reducing cancer and increasing stress resistance. Both of these pathways play a role in the longevity effects of SIRT1. Unfortunately, SIRT1 expression declines with aging. As a result, all of these cancer prevention mechanisms and these stress resistant mechanisms decline with aging.
references:
1. Xiaolei Qiu, Katharine V. Brown, Yehu Moran, Danica Chen, Sirtuin regulation in Calorie Restriction, Biochimica et Biophysica Acta (BBA) – Proteins and Proteomics, Volume 1804, Issue 8, August 2010, Pages 1576–1583
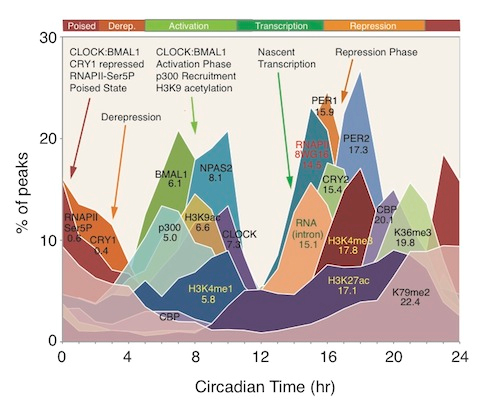
Whereas Sir2 appears to play the role of “gene silencer” in Baker’s yeast, in humans gene silencing is done primarily by SIRT1, SIRT2, SIRT3, and SIRT6. Each of these SIRTs have other functions besides gene silencing, however. They are “redox sensitive” enzymes, like all of the Sirtuins. Since SIRT2 and SIRT6 are two good examples of the two primary “silencing signatures” seen on histones, I will limit the discussion of gene silencing to these two good example Sirtuins.
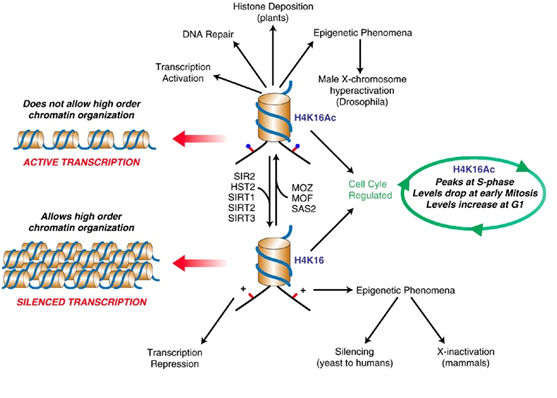
SIRT2 – The gene silencer with the “H4K16 silencing signature”
SIRT2 is a Class III Histone deacetylase. Like SIRT1, SIRT2 has many targets for deacetylation in humans, including FOXO1, p53, FOXO3, alpha-tubulin, and Histones. SIRT2 deacetylates the K16 lysine on the tail of histone protein 4. The acetylation status of lysine 16 on Histone H4 has a critical role in multiple functions in chromatin regulation. Acetylation of H4K16 is associated with active transcription and open chromatin. H4K16 deacetylation is an important silencing mechanism that appears to be “silencing signature” common to SIRT1, SIRT2, and SIRT3. However Here is a diagram illustrating how this works.
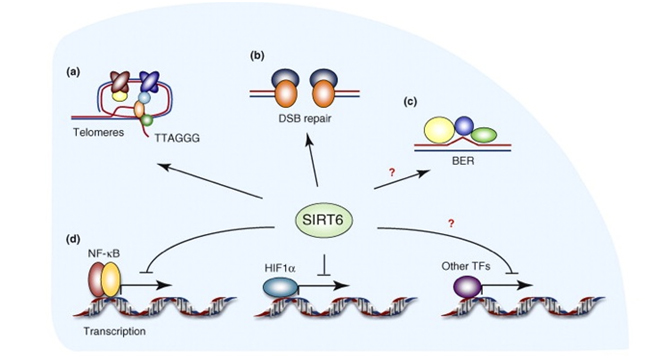
SIRT6 – The ‘Split End Repair” and the “gene silencing signatures” of SIRT6
Whereas SIRT2 was primarily involved with transcription factor deacetylation and H4K16 mediated gene silencing, SIRT6 appears to have multiple roles in maintaining telomere integrity, preventing telomere fusions, and DNA repair. It does have a secondary role in in gene silencing via two other unique “silencing signatures”, but the other functions appear to be more important than gene silencing. Here is a diagram illustrating the main functions of SIRT6.
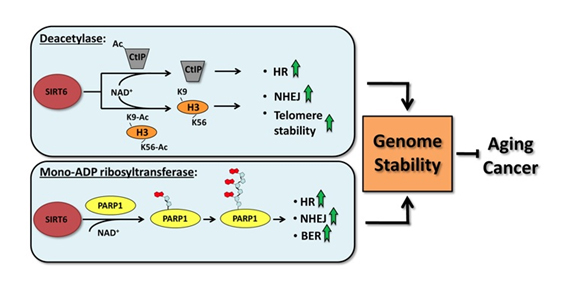
SIRT6 is both a protein deacetylator and a ADP ribosylator. Both functions of the SIRT6 enzyme are very important for genome stability. The diagram below illustrates how this works:
As a deacetylator, SIRT6 has three unique “silencing signatures”. SIRT6 deacetylates two specific lysine side chains on the H3 histone tail – the K56 lysine and the K9 lysine. For this reason, H3K9 or H3K56 deacetylation is a tell tail “sign” that SIRT6 has come by. Also, SIRT6 deacetylates CtIP.
The most dangerous event for genomic stability is a double-stranded DNA break. Unrepaired DNA leads to irregular gene expression, cell cycle arrest, apoptosis, and cancer. To ensure that there are two “redundant” systems for DNA repair, two separate repair systems have evolved – homologous recombination (HR) and non-homologous end joining (NHEJ). The first evidence that SIRT6 was involved in double-stranded DNA breaks came from knockout mice experiments. These mice exhibited a very high incidence of chromosomal rearrangements and breaks. They were very sensitive to ionizing radiation, especially gamma radiation. Now there is good evidence that SIRT6 specifically recruited to break sites after DNA damage. SIRT6 functions in NHEJ by stabilizing the NHEJ haloenzyme, DNA-PK at the sites of the double stranded breaks. SIRT6 also promotes HR by deacetylation of the end resection protein, CtIP. Overexpression of SIRT6 stimulates DSB repair through both the HR and NHEJ pathways by 3-fold. SIRT6 does this by its ADP-ribosylation function. SIRT6 ribosylates the upstream DSB repair factor, PARP1, at lysine 521, thereby stimulating its poly-ADP ribosylation activity. PARP1 facilitates the recruitment of the MRN complex to double stranded breaks, plays a role in the activation of ATM and helps to direct the choice between the NHEJ and HR repair pathways. Additionally, PARP1 is required to promote a non-canonical form of NHEJ (Alternate NHEJ pathway). By this mechanism, SIRT6 can stimulate NHEJ in the absence of DNA-PK.
Stress Preconditioning before you drink your Wine – evidence for a hormetic ROS dose preconditioning for DNA repair.
An interesting phenomena was found by a group at the University of Rochester recently. They were studying the effects of SIRT6 on DNA Strikingly, when cells were pretreated with oxidative stressors prior to overexpression of SIRT6, DSB repair efficiency was massively stimulated by up to 16-fold, suggesting that SIRT6 specifically integrates stress signaling to prime the DNA repair machinery in response to oxidative stress. Not only did ROS preconditioning increase the intensity of the double-stranded break repair response, it recruited SIRT6 to the site of the double stranded break much sooner (30 minute recruitment time vs 8 hour recruitment time). This means that some stress preconditioning before exposure to DNA damage (for instance, chemotherapy, X-rays, or cigarette smoke) will increase the speed and intensity of the DNA repair response by SIRT6. It appears that that both the deacetylation function and the mono-ADP ribosylation function of SIRT6 is needed for this stress response. It appears that both SIRT6 and SIRT1 localizes to double-stranded DNA breaks in response to DNA damage.
The second “song of silence” referred to in the title of this blog is via polycomb group proteins which can silence genes in plants, insects and animals. “Polycomb proteins play a key role in developmental gene regulation in most multicellular organisms, from plants to mammals. They were first defined in Drosophila as factors which allow cells to ‘remember’ patterns of gene expression through successive cell divisions, providing a memory of the cell fate or identity established through differentiation and development. Polycomb proteins play a key role in developmental gene regulation in most multicellular organisms, from plants to mammals. They were first defined in Drosophila as factors which allow cells to ‘remember’ patterns of gene expression through successive cell divisions, providing a memory of the cell fate or identity established through differentiation and development. In mammals, polycomb proteins have important roles throughout an organism’s life. In embryonic stem (ES) cells, they play a crucial role in restraining the cells from going down differentiation pathways, targeting around 1500 genes. They are vital in the establishment and maintenance of different cell types. Polycomb proteins are also found on the inactive X chromosome. — Polycomb proteins function in multiprotein complexes which are recruited to and silence selected genes. They accomplish silencing by chemically modifying the histone proteins around which the DNA is entwined. — Polycomb complexes exist in two forms – PRC1 and PRC2. PRC2 loads onto the chromatin first, modifying a lysine on one of the resident histone proteins by adding methylation groups – H3K27me3. This methylated lysine is recognised by a protein called CBX in the PRC1 complex. PRC1 has a different chromatin modifying activity known as ubiquitylation. Bringing PRC1 onto the site leads to ubiquitylation of another histone protein, H2A. It is this modification that inhibits transcription on the genes located at that site, possibly by blocking the enzyme which transcribes the DNA into RNA(ref).” Reference RYBP-PRC1 Complexes Mediate H2A Ubiquitylation at Polycomb Target Sites Independently of PRC2 and H3K27me3.
A number of publications have focused on the gene-silencing actions of polycomb group proteins, for example, Silencing by plant Polycomb-group genes requires dispersed trimethylation of histone H3 at lysine 27, “In plants and animals, Polycomb-group (Pc-G) genes mediate mitotically stable repression of targets such as homeotic genes that are critical for developmental patterning and growth control. An antagonistic group of proteins, the trithorax-group (trx-G), act as activators. Both groups act to maintain on/off patterns of transcription defined early in development, rather than to set up these patterns, and are important for maintaining cell fates. Although the Pc-G and trx-G have long been thought to cause epigenetic changes in chromatin structure, due to the stable but ultimately reversible alterations in gene activity they promote, the mechanistic basis for their activity has been mysterious. Recent studies have implicated histone modifications as an important component of epigenetic changes. A variety of modifications on the amino-tails of histones have been characterised, of which methylation of lysine residues has been thought to be particularly stable (Jenuwein and Allis, 2001; Fischle et al, 2003). The consequence of lysine methylation can differ both according to which lysine residue is modified and also as to how many methyl groups are added—lysine residues can be mono-, di- or trimethylated (Bannister et al, 2002). For example, methylation of histone H3 at lysine 9 (H3K9) or lysine 27 (H3K27) is generally correlated with transcriptional repression, whereas methylation at lysine 4 (H3K4) is predominantly associated with transcriptional activity (Jenuwein and Allis, 2001; Peters et al, 2003; Ringrose and Paro, 2004). In addition, the level of methylation is important, for example H3K9me3 shows a different distribution from H3K9me1 and H3K9me2 in mammals (Peters et al, 2003). Several enzymes with histone lysine demethylase activity have now been identified, indicating that methylation can be rapidly reversed (Shi et al, 2004; Tsukada et al, 2006).”
Many publications focus on the roles of polycomb group proteins in cancer. For example, Polycomb group protein gene silencing, non-coding RNA, stem cells, and cancer (2011). “Epigenetic programming is an important facet of biology, controlling gene expression patterns and the choice between developmental pathways. The Polycomb group proteins (PcGs) silence gene expression, allowing cells to both acquire and maintain identity. PcG silencing is important for stemness, X chromosome inactivation (XCI), genomic imprinting, and the abnormally silenced genes in cancers. Stem and cancer cells commonly share gene expression patterns, regulatory mechanisms, and signalling pathways. Many microRNA species have oncogenic or tumor suppressor activity, and disruptions in these networks are common in cancer; however, long non-coding (nc)RNA species are also important. Many of these directly guide PcG deposition and gene silencing at the HOX locus, during XCI, and in examples of genomic imprinting. Since inappropriate HOX expression and loss of genomic imprinting are hallmarks of cancer, disruption of long ncRNA-mediated PcG silencing likely has a role in oncogenesis. Aberrant silencing of coding and non-coding loci is critical for both the genesis and progression of cancers. In addition, PcGs are commonly abnormally overexpressed years prior to cancer pathology, making early PcG targeted therapy an option to reverse tumor formation, someday replacing the blunt instrument of eradication in the cancer therapy arsenal.”
Other recent publications relating PcG silencing to cancer include:
There is a significant literature related to polycomb group silencing. Topics include signaling in cancers, signaling in plant, insect and human development, and details of the silencing process. 2013 publications include:
Site-specificsilencingof regulatory elements as a mechanism of X inactivation
DMNT3 GENE SILENCING
The third “song of silence” referred to in this blog title in epigenetic gene silencing brought about by DNMT3 methyltransferases. This subject has already been discussed in detail in the previous blog entry PART 2: Slaying Two Dragons with One Hail of Stones: The Silencing Of Good Genes In Aging And Cancer. “Gene silencing by epigenetic mechanisms is an ordered series of events that normally starts with the methylation of cytosine residues. Scientists studying gene silencing have observed two distinct kinds of CpG methylation: one that is age-related that occurs in all tissues as a function of aging, and a cancer-related pattern of methylation that only occurs in cancerous cells. It is important to point out that with aging, there is a global loss of methylation at sites other than CpG islands. The hypermethylation of CpG islands is the exception to this universal decline in DNA methylation with aging. This can be explained in part by the fact that there are different DNA methyltransferases for these different regions of the genome. DNMT1 is responsible for the maintenance of hemimethylated DNA, whereas DNMT3a and 3b are responsible for the methylation of unmethylated DNA. For this reason, it appears that DNMT3a and 3b are responsible for gene silencing.”
PART B
A masterlist of drugs and natural compounds for cancer chemoprevention
You may download such a list as a Word document by clicking on link below.
Note on bioavailability of plant polyphenols and nano-particle delivery
This blog post touches on biological activities and health benefits of certain plant polyphenols (e.g. Resveratrol) as do a great many other postings over the years. It should be pointed out that for many plant polyphenols, biological impacts observed in vitro may not be fully realizable in vivo based on normal oral ingestion. There are several reasons for that. 1. Chemical structures can be altered by gut bacteria. Sometimes the gut biome helps us in this regard, an example being gut actions on the polyphenols in chocolate (protoanthocyanadins). But in states of altered gut micro biome, we can’t always count on the bacteria. 2. Poor absorption. This is a major issue with many of the polyphenols, especially curcumin. 3. Metabolism by the liver via the first pass effect, this destroys a lot of the polyphenols. The metabolites are not as good signaling compounds. 4. Glycosylation. This makes substances more water soluble, but then they are secreted faster. Also, glycosylation reduces the ability of a substance to become part of the mitochondrial membrane and work as a hydrophobic mitochondrial specific antioxidant. (I am now convinced that the chloroplast outer membranes are places where stress-defensive phytochemicals migrate to in plant cells. In us humans on the other hand, the mitochondrial membranes are where we can control excess baseline ROS leak that occurs in old mitochondria.) 5. Last of all, plant polyphenols tend to be very prone to oxidation. This means they are inactivated with heat (instantly) in water (4 hrs), and in the presence of oxygen, especially with cation catalysts like unliganded iron or copper.
Indeed, poor bioavailability is probably the major reason why clinical trials of certain phyto- substances like resveratrol and curcumin against disease indications like cancers and Alzheimer’s disease have not produced positive results.
We believe there is an elegant solution on the horizon to this issue of poor bioavailability of health-producing phyto substances. That is, the use of nanoparticle polyphenols. Very recently, there has been significant research related to nano-particle delivery of drugs and phyto substances. Because of the importance of the bioavailability issue and the emerging importance of nano particle delivery, we plan to produce a blog on the bioavailability issue and on nano particle delivery in the near future.
DATA DISCLAIMER:
The tables and compilations of data in this and the other blog entries in the Two Dragons series are intended to be illustrative of the main points made in the blog entries. The data are compiled from various sources, in most cases are incomplete, and may contain occasional errors.
MEDICAL DISCLAIMER
FROM TIME TO TIME, THIS BLOG DISCUSSES DISEASE PROCESSES. THE INTENTION OF THOSE DISCUSSIONS IS TO CONVEY CURRENT RESEARCH FINDINGS AND OPINIONS, NOT TO GIVE MEDICAL ADVICE. THE INFORMATION IN POSTS IN THIS BLOG IS NOT A SUBSTITUTE FOR A LICENSED PHYSICIAN’S MEDICAL ADVICE. IF ANY ADVICE, OPINIONS, OR INSTRUCTIONS HEREIN CONFLICT WITH THAT OF A TREATING LICENSED PHYSICIAN, DEFER TO THE OPINION OF THE PHYSICIAN. THIS INFORMATION IS INTENDED FOR PEOPLE IN GOOD HEALTH. IT IS THE READER’S RESPONSIBILITY TO KNOW HIS OR HER MEDICAL HISTORY AND ENSURE THAT ACTIONS OR SUPPLEMENTS HE OR SHE TAKES DO NOT CREATE AN ADVERSE REACTION
A collection of stories and explanations by Jim Watson and Vince Giuliano
The stories are fantasies about new kinds of bars that could possibly exist in the future. The Hormesis Bars are ones where all the drinks produce stresses. According to what many believe, these stresses could actually make you live longer and more healthier. This is because most of the drinks served in the bars consist of deadly poisons or toxic actions. The stories are heavily interspersed with explanations on the science behind the purported health effects of the drinks and with links to other blog entries and citations.
Just like for the amazing stunts that appear commonly in TV, we have to warn you DO NOT TRY THESE DRINKS OR TREATMENTS YOURSELF. If such bars were to exist in the future, one thing is for certain, and that is that the doses will have very carefully been worked out so as to produce safe hormetic results. In most cases we do not now know what those doses should be. And, appropriate doses could depend on the individual and their momentary state. And yes, wrong doses could lead to disastrous results. Hormetic bars would probably be reguIated by very strict laws.
These stories are about various ways of realizing health via hormesis, so if you don’t know what that is we strongly suggest you find out before trying to understand the implications of drinking drinks from the hormesis bars. A good initial approach could be to view our PowerPoint presentations downloadable from this link. And you can find many other blog entries related to hormesis in this blog such as Multifactorial hormesis – the theory and practice of maintaining health and longevity. And, we suggest other citation links throughout the stories.
The Hormesis Bar Menu
Imagine a large stone building with an ornate classical façade decorated with stone gargoyles in various states of suffering. There is a desk and a gate at the entrance, and in order to get in you have to sign a 9-page fine-print document that absolves the bar operators from any legal liability for what might happen to you as a result of your visit. Inside there are is a collection of shops – the individual hormesis bars. Most are dark and exotic with rich marble floors, mahogany walls and lots of polished brass. Some seem to be filled with elaborate plumbing.
Welcome to the Hormesis Bars. We serve “stress drinks” all day and recommend resting at night to recover. All of our “stress drinks” are served in small shot glasses. A big glass of these drinks would kill you. Many believe that these “small shot glass stress drinks” are bad for you, but we believe they will actually make you live longer and more healthier. Just think of them as being ”stress chasers”…….small shots of stressors that are too small to kill you, but big enough to vaccinate you from getting killed from a 24-once glass stress, such as a heart attack, a stroke, a big dose of nuclear radiation or getting hit by a bus.
We think our drinks do much more than chase away the liberal, free radicals. They actually create stress signaling molecules in your body within minutes or hours, but several may protect you from larger stressors for weeks to months. This idea of a “small shot glass of stress” before a “super sized stress glass” is called preconditioning.
Our Famous Seven Sirtuin Silencing Spices of Life
Near the entrance is a special kind of spice shop. We have discovered seven spices that make your drinks taste or work better. We call these the “Seven Sirtuin Silencing Spices”, although we are not exactly sure what they are silencing. If you sprinkle some ”Sirtuin spice” on your main menu item from one of the hormesis bar menus, the Xenohormesis Bar menu, or the Xenobiotic Bar menu, we think that your drink will give you a bigger “shot in the pants”. It may help with the tastes as well, since some of our menu items are a “bitter pill to swallow”, especially the ones on our Xenohormesis Bar menu. All of the Sirtuin spices will make your drink taste better, kill cancer, and keep you alive. We now offer 7 flavors of Sirtuins which well call the “spice of life” in that they will make you live longer. Here are our menu offerings “du jour”. We have made some suggests as to what menu items you should add each spice to. Enjoy the bar!
The Sirtuin spices are our most popular spices that activate stress coping pathways, making them work better. These spices also silence bad genes, help get rid of ammonia, help with sugary diets, and make the microtubule railway transport things faster. We offer 5 varieties that deacetylate and 2 varieties that ADP-ribosylate.
SIRT1 – this spice is good for making the stressor more efficient at turning on your stress coping genes. It goes well with the following menu items (transcription factors): FOXO3a, p53, Nrf2, NF-kB, PARP-1, Ku70, Notch. It also goes well with the following combo menu items (co-activators): PGC-1a, p300, NCoR. We have talked about this spice in the three most-recent blog entries (ref)(ref)(ref), and in several others.
SIRT2 - this spice is good for suppressing bitter tasting DNA, such as non-coding repetitive DNA that has no replicative value and should be silenced, you know, like messy retrotransposons that love to duplicate themselves crowding-up our genomes. SIRT2 is good at silencing this at Histone 4 at lysine #16, by removing the acetyl group there (H4K16)(ref). This keeps your genes stable(ref). This spice also goes great with our FOXO1 menu item to help you eat yourself for dinner (autophagy). This spice also goes well with our FOXO3 and p53 menu items to help you become resistant to cell death (apoptosis) and resist those liberal, dangerous, free radicals (ROS). (ref) This spice will also allow you to get around better by removing acetyl groups from your alpha-tubulerailway transport system. You can get important stuff from point A to point B better this way on your little intra-cellular railroads(ref).
SIRT3 - this spice goes well with anything from our mitochondrial menu items(ref)(ref)(ref)(ref). This spice variety goes well with PGC-1a and with UCP-1. If you are a long distance runner, we suggest PGC-1a for turning on your PPAR gene. (ref)(ref). If you are cold all the time, we would add SIRT3 and UCP1 (ref) together to keep you warm. We also suggest you consider adding this spice to SOD2, a popular mitochondrial menu item that will help you deal with rogue, free radicals that leak out of the electron transport system. We have heard reports that the leak rates have been rising from 2% to as high as 40% in some cases, due to the bad local energy economy (in certain mitochondria) (ref).
SIRT4 - this spice goes well with anything from our mitochondrial menu as well, but is very unique(ref). It has a very different flavor than the other spices above, but is similar to our SIRT6 spice below. These spices do not take away acetyl groups, but adds an ADP-ribosyl to your menu item. This spice goes great with sugary menu items, since it stimulates insulin secretion.
SIRT5 - this spice also goes well with our mitochondrial menu item, and is our best spice for getting rid of ammonia. It gets rid of ammonia, turning it into urea which allows you to “piss away your ammonia”, rather than making you sick.(ref)(ref)
SIRT6 - this spice is similar to our SIRT4 spice above in that it does not take away acetyl groups, but adds ADP-ribosyl to any menu item. This spice is our best survival spice, in that it helps you repair your DNA, protects you from death (apoptosis) and helps you survive. Unfortunately, it stimulates inflammation to do this (by promoting NF-kB and TNF, the opposite of SIRT1 in this respect).(ref)(ref)(ref)
SIRT7 - this is our favorite spice, since it controls protein synthesis in the cell nucleus. This spice deacetylates nuclear histones, silencing genes for protein biosynthesis.(ref)
The bars are grouped into three clusters: Hormesis Bars, Xenohormesis Bars, and Xenobiotic Bars.
I. The Hormesis Bars -Legendary stories, legendary drinks
These drinks are “the real thing”, not some fake namby-pamby “mimetic” drink. (There are no tiny umbrellas or slices of lemon on these drinks….just hard core, punishing drinks that will make a grown man cry and say “Uncle”). Most of the bars serve drinks are also natural and are ”xenofree,” exceptions being our Xenohormetic Bar and Xenobiotic Bar below. (Not that the xenohormesis or xenobiotic drinks are bad……they are just not natural.) Unfortunately, these drinks will make you sweat, shiver, wither, or gasp for air. Some are not for the faint hearted (especially for those visiting the Radiation Bar or the Igloo Bar). We recommend mixing drinks. A good mix is combining a drink from the Desert Bar and the Igloo Bar. Another good combination is the Starvation Bar and the Hypoxia Bar, but few can take this “mixed drink” without passing out. We have included some amazing stories below that can be told while sitting on the bar stools. These stories are hard to believe, but no one questions their legendary status as “heroic fables”. (They are much more believable than fisherman’s stories about catching big fish).
A. The Oxygen Bankruptcy bar - Bar story: The legend of the bartender at the Oxygen Bar who ran out of oxygen
This our most popular bar in the plain-hormesis section. It is one of our two “gasping bars” where clients are gasping for a breath of air. (The other is the Hypoxia Bar). It involves extreme physical exertion, going past the point of sweating and into the zone of oxygen debt. Fast treadmills, powered staircases and other fiendish machines force the exertion. A “drink” consists of a timed ride on such a machine. All drinks at the oxygen bankruptcy bar cause pain. They all stimulate a lot of Nrf2 and AMPK. (ref)(ref)(ref)(ref)(ref). If you have too much mTOR or NF-kB in your system, these drinks will get rid of mTOR and NF-kB(ref)(ref). After the pain, you will feel better. You have to wear a cardiogram monitor while on a machine and it will shut down automatically if there is any sign of an impending heart attack.
Legend has it that a clever bartender at an “Oxygen bar” ran out of oxygen on day. There were a lot of customers hanging out so he started selling drinks with low oxygen, instead of high oxygen. He hoped nobody would notice. But to his surprise, he found out that low oxygen levels were actually better for his clients than extra oxygen. So, he changed the name of the bar to the “Oxygen Bankruptcy Bar” and raised the price of drinks. This improved his profit margin a lot since his cost for the oxygen ingredient went down to zero. Later he found out that he did not have to lower the oxygen, but could just make his clients exercise vigorously while breathing ordinary air. So he installed heavy exercise machines. The exercise had to be so vigorous that the clients were in pain. He called this concept the ”no pain, no gain” principle and went on to start a gym called Gild’s Gym. The claim is that if you do not have a sour taste in your mouth or if your muscles are not burning, you have not exercised hard enough. The pain is due to lactic acid, and is actually NOT ”oxygen bankruptcy”. In this sense, the term “Oxygen Bankruptcy bar” is not very accurate either. The true “oxygen bankruptcy bar” is actually the Hypoxia bar.” (see below). .
B. The Desert Bar – also known as the Heat Shock Bar -
This bar consists of hot sauna rooms, hot tubs, sweat lodges and infra-red chambers, places that can make you hot. There is also a “light treatment” area where you can put on clothes with circulating hot water.
Bar story: The legend of the origin of the term “shock and awe.” This bar is one of our two “shock bars”. Both produce “shock proteins” which can help you survive any “shock and awe”. This “shock bar” makes a type of protein called “heat shock factor”, or HSF. There are 4 HSFs, named HSF1, HSF2, HSF3, and HSF4. (ref)(ref)(ref)(ref) The legend of the name of the “heat shock bar” comes from the US Military. Troops were training in hot weather for the Iraq invasion and were “shocked” at how hot it was outside in Kuwait (it was about 42-45 degrees C). To the surprise of the Army generals, the soldiers who survived these extremely hot conditions became stronger. To memorialize these “heros of heat”, the US Military called the Iraqi Invasion the “shock and awe” attack. The “shock” was the hot weather that the soldiers endured and the “awe” was the amazement of the big fat generals in their air conditioned HumVees, who did not do as well as the soldiers in the heat. Now we know that the ”heat shock” survival advantage of the solders was due to the “Heat Shock proteins” produced by the desert sun heat. (Not everyone believes this legendary version of the story, but it is still a great story to tell sitting on a stool in an air-conditioned bar.) The chaperones at this bar are very good refolding proteins, getting our clients back into shape, if they become unfolded. So, this bar is very popular with our “unfolded protein” customers, since we offer chaperones for any client who may become disheveled with the heat. Expression of such chaperone proteins is called the “unfolded protein response.” We have chaperones who specialize in refolding in all 4 rooms of the bar (mitochondria, ER, cytoplasm, and nucleus).
All of our ”shock drinks” at this bar are served hot. They can be steamed, served dry, served in a tub, or served in a sweat house. We suggest drinking this at 40-42 degrees C and not drinking for more than 20-30 minutes or you may pass out. If you are sweating profusely and are begging to go to the Igloo bar, then you have had enough. We recommend ”bar hopping” from the heat shock bar to the Igloo bar and then repeating this over and over.
C. The Igloo bar- Bar story: The legend of “Ig the Explorer” and the true story of the ”200 club”
This is the other “shock bar.” It consists of refrigerated rooms and lockers, icy water tubs, actual igloos and powerful air-conditioners that can blow on you. Clothing here is frowned upon. One room has a thick layer of snow on the floor and you are encouraged to lie down there and make snow angels,
Many of our clients come here after visiting the Heat Shock bar, but then start “bar hopping” back and forth from the Heat Shock Bar to the Igloo bar. This type of “bar hopping” is very popular with our guests from Scandinavia. They claim it is really good for the immune system, but we don’t know exactly how this works. We encourage, but do not mandate the Scandinavian dress code at this bar, however (In Norway, Sweden, Finland, and Denmark, they don’t wear any clothes!). The original Igloo bar was not in Scandinavia, but was built by prehistoric explorers who crossed the Bering Strait on their way to the “New World” from Asia. Back then, the Bering Straight was mostly land and only had a few shallow water areas covered with ice. Legend has it that one of these explorers had some Neanderthal heritage and was not very smart. His name was ‘Ig” (this is the origin of the word ignorant). Ig ventured out onto thin ice and fell into the frigid waters of the Bearing Straight (which was very shallow back then). Despite being really stupid, the “cold shock” stimulated Ig enough to climb out of the ice cold water. Instead of dying, he survived and became stronger than his (smarter, non-Neanderthal) peers. Later during the expedition, a largeGrizzly bear attacked the expedition. Due to all of the “cold shock proteins”, Ig was able to fend off the Grizzly bear and save the lives of all of the (smarter, but weaker) expedition members. To memorialize the heroism of “Ig the Explorer“ and his miraculous survival from falling through the ice, the explorers built a house out of ice and named it “Igloo” for “Ig the explorer”. This legend has survived many winters and has taken on “fable status” in communities near the North Pole, Not to be outdone by North Pole legends, the South Pole Scientists have created their own bar with legendary stories. They call this bar the “200 club”. Here South Pole stress aficionados sit in a hot tub at 110 degrees F, then run naked outside into the frigid outdoor temperatures when it is -90 degrees below zero, F. Then they run back into the hot tub, going through a temperature shift of 200 degrees F. This sounds extreme, but the scientists at the South Pole get very bored with nothing else to do. They now are making up extreme stories and legends to compete with the legend of “Ig the Explorer”. These scientists are not very creative, however, since they have very few genes left from Neanderthals, which “Ig the Explorer” had in his genome. The cold-shock hermetic response is described in our PowerPoint presentation on Multifactorial Hormesis.
D. The Hypoxia bar- Bar story: The legend of the Sherpa who forgot the oxygen cylinders
This is not a “shock bar”, but is definitely a “gasping bar.” It is a competitor with the Oxygen Bankruptcy Bar, owned by a different franchise-holder. It consists of sealed decompressed-chambers. Or, you can get your hypoxia wearing masks. At the Hypoxia bar, most people would sell their first-born son or daughter for a bottle of oxygen. Unfortunately, there are no oxygen bottles at this bar. Instead, customers are served up only air with 8-15% oxygen, which is the equivalent of climbing a mountain with an elevation between 14,000 feet and 22,000 feet. The hypoxia bar does NOT put you into into “oxygen bankruptcy”, but will make you oxygen insolvent.
Legend has it that a Mountaineering expedition brought along a Sherpa of Neanderthal ancestry. This Sherpa was responsible for packing all of the oxygen cylinders into base camp. He labored with the heavy backpacks, delivering them to base camp in a timely fashion, only to find out that his buddy sherpas had played an old trick on him. (the backpacks were full of rocks! No oxygen cylinders were in the packs! ) This was a disaster, considering the tight time window for the summit climb. The Neanderthal Sherpa was in such great shape from bringing thebackpacks up the mountain that he carried the mountaineers to the top of the mountain in his backpack. From this day on, the concept of climbing a mountain without supplemental oxygen became vogue, thanks to the Neanderthal Sherpa. On Mt. Everest, records indicate that those who climb to the top without oxygen actually have a higher survival rate than those who climb with oxygen. This is probably due to the fact that the climbers who climb without oxygen are in better shape and have acclimatized better. (This part of this legend is true!).
There are obvious negative effects of hypoxia, such as cell death (necrosis and apoptosis) as well as excessive angiogenesis, but transient hypoxia with controlled dosing does not seem to cause these negative effects. Intead, transient low levels of oxygen stimulates cellular housecleaning, including a specific form of autophagy called “mitophagy”(ref)(ref). Mitophagy selectively gets rid of bad mitochondria that are “leaking” those liberal, free radicals out of electron transport. As a result, hypoxia gets rid of the source of most of the free radicals. Hypoxia also gets rid of aggregated proteins that accumulate in cells. (another form of autophagy). Hypoxia is so effective at cellular house cleaning (autophagy) that the Hypoxia bar is now starting a maid service for removing ‘unfolded proteins” that could cause Alzheimer’s disease, Diabetes, Parkinson’s disease, and many other age-related amyloidopathies. The Hypoxia bar can get rid of unfolded proteins that the chaperones at the Heat Shock bar could not refold.
This means that the Hypoxia bar is a better housecleaner than the Heat Shock Bar. To everyone’s surprise, hypoxia also stimulated stem cells to divide and stimulated stem cells to circulate. The effects of hypoxia on autophagy (and mitophagy) as well as the effects on stem cells may be why we should visit the hypoxia bar on a regular basis, to stimulate our stem cells and clean our houses(ref). Hypoxia may be the best stimulus to schedule before you have a heart attack, a stroke, a concussion, or before getting hit by a bus.
E. The Curie Bar- No Bar Stories here! The true story of Ikarus, Greece and the science of stimulating DNA repair pathways
One wall of this bar consists of cozy booths having lead walls, each containing an old-fashioned dental x-ray machine. The “drinks” served in these booths consist of mildly radioactive mineral water accompanied by timed and controlled doses of whole-body x-rays. The drinks served at the bar area on the opposite wall consists radioisotopes with short half lives. For example there is the “Let’s See Margarita” which contains technetium-99m, a radioisotope commonly used for medical diagnostic applications. This radioisotope has a 6-hour half life and has been used for tens of millions of diagnostic procedures.
This bar has been condemned by the entire scientific community as being a “dangerous bar”. This is surprising, however, since there is strong scientific evidence for the molecular mechanisms of how transient “radiation drinks” could promote health. See the blog entry Radiation hormesis . The description below touches on the history of radiation, the molecular mechanisms of action, and the claims from radioactive hot springs from around the world. This bar is named after after Madam Curie, the pioneer in the study of radioactivity, the first woman to win a 1 Nobel prize, the first woman to win two Nobel Prizes, and the first person to win a Nobel Prize in multiple disciplines. Like Madam Curie, the radioactivity she studied may be the first ”hormesis mechanism” that evolved for protecting the “language of life” – DNA. Today we know that low doses of radiation stimulate DNA repair through the activation of four transcription factors, PARP-1, PARP2, ATM and Ku70. (ref) In response to low dose, sublethal radiation (or xenohormetic compounds). PARP-1 and PARP-2 activate two DNA repair pathways called ”Single strand break repair” (SSBR) and one called “Base Excision Repair” (BER). Both of these repair single-stranded DNA breaks. In response to low dose, sublethal radiation (or xenohormetic compounds), Ku-70 activates the DNA repair pathway called “Non-homologous end joining” (NHEJ). Thus the Curie bar can repair your “single-stranded break-ups and your “double-stranded breakups”.
If this is so, then why does it seem that the whole world is against radiation? The key is ”getting the dose right” and the “exposure time right”. With the apparent world-wide opposition to any radiation exposure, is there any evidence from nature or human history for low dose exposure to radiation as being beneficial? The answer is yes, in terms of experimental and empirical evidence, in terms of understanding how radiation hormesis works, and in terms of folk medicine. To discover the folk medicine aspects, you need to visit the following eight locations (with an open mind):
1. The Greek island of Icarus and its radioactive hot springs
2. The Isle of Ischia in the Bay of Naples,
3. The Radium Palace, Czech Republic
4. The Bad Gastein in the Austrian Alps
5. Ischia in Italy
6. The Radium Palace, Czech Republic
7. Fairmont. Springs and Resort, Banf, Alberta
8. The Free Enterprise Radon Health Mine, Bolder, Montana
We will discuss Icarus in a blog entry now under preparation on “Blue Zone” places having populations with extraordinary longevity. Tales From the Nuclear Age traces some of the traditional uses of radioactivity for health purposes. Many of these are controversial or have been dismissed as quackery. Yet, there is a strong scientific underpinning for such benefits being real.
Theoretical therapeutic effects of radiation hot springs - How heat works and how radiation works
Most of the therapeutic effects of the radiation hot springs can be explained by two scientific mechanisms: heat and radiation. Unfortunately, few or no serious scientific studies have been done in the places mentioned relating health benefits to heat or radiation, but here are some well-documented beneficial effects seen in the laboratory:
Heat mechanism of action:
It is likely that the heat works via gene expression and autophagy triggered by the following 4 mechanisms:
1. Cytoplasmic Heat Shock Factor migration into the nucleus, also referred to as the “heat shock response.” This up regulates dozens of genes such as Hsp70 and Hsp90.
2. Endoplasmic reticulum Unfolded Protein Response, which triggers gene expression via 3 mechanisms – IRE-1/XBP, PERK, and ATF6 pathways. This up regulates ER chaperone genes, down regulates protein synthesis, and up regulates protein degradation via proteasomes.
3. Mitochondrial Unfolded Protein Response, which triggers gene expression via a 2-step mechanism: JNK2/Jun pathway activation => expression of CHOP and C-EBPb => expression of mitochondrial chaperone proteins and proteases.
4. Heat directly up regulates the Nrf2/Keap1 pathway via HSF1, which triggers gene expression via the ARE genes which produce antioxidants.
5. Hydrogen sulfide in the air from sulphur hot springs directly up regulates the Nrf2/Keap1 pathway, independent of HSF1. This also up regulates the ARE genes which produce endogenous antioxidants.
6. Autophagy/mitophagy activation via stimulation of all of the above – HSF, ER-UPR, Mito-UPR.
Radiation mechanism of action:
It is likely that the low dose X-ray radiation (XRT) works via gene expression triggered the following mechanisms:
1. XRT directly creates ROS, which then activates Nrf2/Keap1 pathway, which triggers gene expression via the ARE genes.
2. XRT directly triggers PARP-1 and PARP-2 which then activates 3 DNA repair mechanisms:
a) BER – The DNA base excision repair system (BER) which repairs single stranded DNA breaks,
b) SSBR or NER- The single stranded base repair (SSBR) which repairs single stranded DNA breaks, and
c) NHEJ – The double stranded DNA repair system, which is activated by PARP-1, but not PARP-2
THESE PATHWAYS ARE ACTIVATED WITH LOW DOSE XRT !
3. XRT directly triggers Ku70 and Ku80, which then activates the double-stranded DNA repair system called non homologous end joining (NHEJ). This is a distinct NHEJ repair pathway from the one that PARP-1 activates.
4. XRT directly triggers DNA-PKcs. This is a another mechanism or portion of the DNA repair system for double-stranded breaks (NHEJ).
5. XRT triggers Autophagy/mitophagy activation via PARP => AMPK.
If by this time you are wondering whether this whole blog post is a gigantic spoof, just check PubMed.org by searching on “radiation hormesis.” You will find 258 research articles as of this writing.
This is another bar that is very inexpensive to operate, since it serves very sparse California-style plates that contain hardly any food at all. And what is served is as empty of nutrients as possible. The first starvation bar resulted from pure selfishness. Back in ancient Greece,a bar-and-grill proprieter. Καθομιλουμένη wanted to save money to increase his profit margin. So, Καθομιλουμένη started to cut portion sizes. Surprisingly, his patrons, mostly intellectuals and sport-oriented people with some diplomats and philosophers, started to get healthier. And dogs fed Καθομιλουμένη’s tiny portion sizes of dog food lived longer. The word was spread quickly by Pittacus, a patron and one of the seven wise men of ancient Greece, and what came to be known as the Starvation Bar flourished, So, Καθομιλουμένη cut the served portion sizes further and continued to do this until they were nearly gone. Thus was born the first Starvation Bar and practical application of a field known today as “Caloric Restriction” or CR.
Not-eating at this bar activates all of the above-mentioned pathways – SIRT spices, FOXOs, Nrf2, AMPK, mTOR inhibition, autophagy, etc. This is not a “shock bar” either. It is also not a “gasping bar” like the Oxygen Bankruptcy bar or the Hypoxia bar. Today we know that short term starvation or long term calorie restriction is good for you. The survival advantage for this phenomena is CR. CR has been shown work via many different mechanisms, including Insulin/IGF-1 inhibition, Sirtuin activation, FOXO3a activation, mTOR inhibition, Nrf2 activation, AMPK activation, and the activation of autophagy – all key health and longevity matters often discussed in this blog.
A good review of what is known about CR follows here. If you want to dig a bit deeper you can go back and read about it in several blog entries(ref),(ref),(ref). The most recent blog entry PART 3: Slaying Two Dragons with the Sound of Silencecontains a discussion relating Calorie Restriction -> Sir2 Activation -> Longevity.
Lower animal studies of CR
Animal studies in all species except for primates have shown that CR extends lifespan, suppresses tumorigenesis, and prevents obesity. CR suppresses the carcinogenic action of many carcinogens, including polycyclic hydrocarbons, benzo(a)pyrene, DMBA. alkylating agents and methylating agents such as diethylnitrosoamine, aromatic amines, and p-cresidine. CR also suppresses the formation of cancer from radiation. CR also suppresses the formation of spontaneous tumors (no carcinogens or XRT).
Non-human primate studies of CR
The studies in non-human primates have shown longevity benefit in one long study and did not show longevity benefit in another study (both over 20 years of study). Both were very well designed and executed studies. The Wisconsin National Primate Research Center study showed a longevity benefit with 30% CR in Monkeys who were started on CR as young adults between 7-14 years of age. The NIH Center study showed no longevity benefit with young and old onset CR (16-23 yrs of age). Both studies clearly showed a health benefit, however. This included a lowered serum cholesterol, triglycerides, fasting glucose, and plasma-free isoprostanone. CR dramatically reduced the risk of cancer in the young-onset CR group as well. In summary, Monkeys are less likely to develop diabetes, cardiovascular diseases, obesity, autoimmune disease, and cancer with CR, but any effect on longevity is still questionable.
Human Studies
The best evidence for CR in humans comes from the island of Okinawa, Japan. Okinawans consume a much lower number of calories compared to Japanese in the rest of Japan. In a US study of CR over a 6 month period in overweight, non obese humans, CR was very beneficial on two biomarkers. This study showed that a 10% CR with exercise, a 10% CR without exercise, and a 14% CR without exercise all improved two biomarkers of longevity – lower fasting insulin levels and lower body temperature. The Biosphere 2 project involved a 30% CR practice by the 4 men and 4 women who lived in the closed system for two years. Many of the anti aging effects seen in animals were observed in these 8 individuals. The TNO study in the Netherlands involved 16 subjects on a 20% CR regimen and they were compared to 8 ad lib feeding controls. This study also showed health benefits such as decreased fat mass and lowered blood pressure.
How CR works:
CR works via the following mechanisms:
1. Decreased Insulin and IGF-1 IGF-1 has been shown to enhance growth of the following tumors: prostate, bladder, breast lung, colon, stomach, esophagus, liver, pancreas, kidney, thyroid, brain, ovarian, cervical, and endometrial cancer cell lines. The mechanism of action for this is that IGF-1 induces the phosphorylation of p53, which makes it migrate to the cytoplasm, thereby preventing or suppressing apoptosis. Mice with disruptions of the IGF-1 pathway live 20-55% longer than wild type mice. Mice with over expression of IGF-1 via the keratin-5 promoter exhibit increased spontaneous tumor development and susceptibility to carcinogens. In humans, elevated levels of IGF-1 are associated with premenopausal breast cancer, and an increased risk of developing prostate cancer. Unfortunately, although there are clearly some benefits of declining IGF-1 levels with CR, there are some side effects of declining IGF-1 levels. The effects of low IGF-1 include muscle wasting and declining mobility. Another major side effect of declining IGF-1 levels is osteoporosis.
2. Increased tolerance of oxidative stress – Nrf2 and FOXO - CR definitely increases ROS stress tolerance. This may be primarily due to an increase in antioxidant enzymes via an Nrf2/Keap1 mediated pathway and the FOXO pathway. Both of these pathways control different genes involved with antioxidant enzyme expression. There is also some evidence that CR also reduces baseline ROS and baseline RNS. The increase in antioxidant enzymes as well as the decrease in ROS/RNS explain the increased tolerance to oxidative stress seen with CR.
3. Increasing serum corticosterone - CR has been shown to increase serum corticosterone and urinary excretion of corticosterone metabolites. Glucocorticoids induce apoptosis and reduce inflammation in a variety of tissues. This is probably another mechanism of action of CR. On the other hand glucocorticoids promote inflammation so the mechanism here may not on the whole be a positive one.
4. Decreased leptin - Leptin is a hormone that is secreted by fat that regulates appetite, appetite control, and energy metabolism via hypothalamic influence. Leptin is a proinflammatory cytokine and may be why there is increased levels of inflammation with obesity. Leptin decreases with CR even more than IGF-1 decreases with CR. Higher leptin levels correlate with prostate cancer risk.
5. Increased glucagon - CR also affects another hormone – glucagon. Glucagon levels increase in response to CR. This is mediated through the FOXA transcription factor, which is different from the FOXO3a transcription factor.
6. Cytoplasmic SIRT1 effects. - Studies of the plant polyphenol have suggested that another mechanism of action of CR is via the activation of NAD+ deacetylases called Sirtuins, substances we have already discussed above Although there are 7 types of Sirtuins, they are all dependent on NAD+ and either deacetylate proteins or add ADP-ribosyl groups to proteins. SIRT1 is the most well understood deacetylase of this family. It appears that CR activates this enzyme as well as several of the other Sirtuins. The downstream effects of SIRT1 activation include driving a dozen transcription factors into the nucleus, including Nrf2, FOXO3a, Ku70, PARP-1, p53, Notch, and NF-kB.
7. mTOR inhibition - CR also works via the inhibition of the cytoplasmic energy sensor, mTOR. mTOR inhibition also induces autophagy and this may be the primary mechanism by which CR works via this pathway.
8. Mitochondrial SIRT3, 4, and 5 effects - There are 3 mitochondrial SIRTs that deacetylate about 400 mitochondrial proteins (30% of the total of 1,500+). They are SIRT3, SIRT4 and SIRT5. These 3 Sirtuins deacetylate the proteins in the mitochondria, making the mitochondria more efficient. The other beneficial effect of Sirtuin activation by CR is the induction of autophagy. For mitochondrial, this means “mitophagy”.
9. TFAM up regulation- CR induces the transcription factor TFAM which is responsible for gene expression for all of the nuclear encoded mitochondrial proteins. TFAM is activated by PGC-1a, creating a “3-way mechanism” of how CR is beneficial for mitochondria: SIRT3-5, PGC-1a, and TFAM.
10. PGC-1a activation - CR stimulates the cofactor that is the central regulator of mitochondrial biogenesis, PGC-1apha. This cofactor is vital to producing new, healthy mitochondria by the process of mitochondrial fission.
11. NF-kB inhibition- CR also works by down regulating inflammation. This occurs via Sirtuin deacetylation of NF-kB. There also appears to be a reciprocal arrangement between Nrf2 pathway activation and NF-kB inhibition. This may be a ROS-driven “see-saw” mechanism (i.e. ROS up regulates Nrf2 and down regulates NF-kB).
12. FOXO activatiton - CR also works by FOXO mediated metabolic adaptations and stress resistance induction. The FOXO transcription factor codes for many of the antioxidant enzymes not controlled by Nrf2. FOXO also works hand-in-hand with PGC-1a for increasing mitochondrial production. This mechanism of action may also explain the reductions in baseline ROS seen with CR.
13. Ribosomal biosynthesis - CR up regulates the S6K1 pathway of ribosomal synthesis, a process that involves the inhibition of mTOR upstream.
14. AMPK activation - CR acts to activate AMPK, which is another cytoplasmic redox sensing system. There is mounting evidence that this may be a more-dominant mechanism of action for CR than the Insulin/IGF-1 pathway, the mTOR pathway, or the other pathways listed above. The most important downstream effect of AMPK activation appears to be autophagy.
15. Reciprocal causation between SIRT1 and eNOS activation - CR induces a very interesting “reciprocal causation” between SIRT1 and eNOS. CR stimulates endothelial nitric oxide synthesis of nitric oxide by SIRT1 deacetylation of eNOS. eNOS activates the promoter for SIRT1. This SIRT1 activates eNOS and eNOS activates SIRT1. The net effect is more nitric oxide synthesis and more protein deacetylation.
16. Prevention of Gene silencing by DNMT3a – 5-mC hypermethylation of promoter sites - Alterations in gene expression occur with aging and appear to be mediated by epigenetic mechanisms that are dependent and independent on Sirtuins. Three hallmarks of aging seen in the nucleus incude the decline in the Lamin scaffolding system that lies just inside the inner nuclear membrane. This appears to be a major cause of aging in the accelerated aging diseases such as progeroid syndromes, especially Hutchinson-Gilford syndrome. Another hall mark is the gradual decline in histones and spermidine, which is thought to be a function of the gradual increase in heterochromatin formation and the compaction of the silenced DNA. The most intriguing feature of aging in the nucleus is the epigenetic changes that occur before heterochormatization occurs. This includes global demethylation of non-promoter CpG residues, as well as focal hypermethylation of CpG islands within the promoter sites of important genes that are supposed to be transcriptionally active. Hypermethylation of CpG islands is clearly associated with a reduced gene expession and eventually gene silencing. CR prevents or delays this phenomena through mechanisms that are independent from Sirtuins and from ROS or antioxidant genome protection by molecules like SOD. In other words, CR has a direct effect on maintaining gene expression and the prevention of promoter site CpG hypermethylation that occurs with aging.
17. Prevention of Gene activation by TET – 5-hmC (hydroxymethycytosine) prevention of gene activation in euchromatin.
In the brain hippocampus, a specific pattern of DNA methylation occurs with aging. Whereas GENE SILENCING by promoter site hypermethylation with 5-methylcytidine is the classic method of gene silencing, in the brain, hydroxymethylcytosine is a different form of GENE ACTIVATION that occurs as a secondary modification of 5-methylcytidine. Specifically, a family of enzymes called ten-eleven translocation enzymes (TET) produce a secondary marker of gene silencing called hydroxymethylcytosine (5-hmC). 5-hmC has a lower affinity to methyl binding proteins and is associated with chromatin opening, rather than chromatin closing. 5-hmC is associated with gene activation, rather than gene silencing.
18. Prevention of Epigenetic Drift - There is evidence that the epigenetic markers of the epigenome vary with aging. Genes that are supposed to be silenced are activated, such as repetitive DNA. Genes that are supposed to be active are silenced, such as tumor suppressor genes. This phenomena is age-dependent and affects the epigenetic patterns of about 15% of the total genome, which includes both non-coding DNA and coding DNA. CR has been shown to decrease epigenetic drift.
19. Prevention of Telomere shortening - Telomeres also shorten with aging. CR has been shown to slow the decline in telomere shortening through a phenomena that is not dependent on replication or cell division. Specifically, this is a mechanism that appears to be more associated with oxidative stress inhibition of telomere shortening.
20. Improve Hormesis with CR (via H2O2)- Hormesis is defined as a dose-response phenomenon characterized by low-dose stimulation and high-dose inhibition. There is a large body of evidence that hormetic pathways are cytoprotective and play a role in health and the prevention of disease. (ref)(ref) This includes the Nrf2/ARE response elements, the FOXO pathway, autophagy, AMPK, mTOR, and many other pathways involved with the cellular adaptive response to stress. Unfortunately these hormetic adaptive responses decline with aging. There is some evidence that CR can halt or delay the decline in adaptive responses to stress. The evidence is not that strong yet for the effects of CR, however. The best study for this showed an increase in longevity in a catalase-deficient yeast model, where the catalase-deficient yeast produced more H2O2. The net effect of this was NOT a decrease in lifespan, but a longer lifespan. The increased H2O2 appears to have a longevity effect via the activation of p53, NF-kB, AP-1, and other transcription factors. In summary, higher levels of H2O2 resulted in lower levels of superoxide anions via a hormetic, adaptive response where SOD is induced. This is one of the best pieces of evidence for hormesis to date.
Clearly many of the 20 mechanisms just described can impact each other in complex feedback loops and are not independent, Some, like telomere shortening, might best be characterized as “downstream” effects of others.
CR Mimetics
One corner of the Starvation Bar is for eaters who really like eating but still want the benefits of CR. So,they want to try something else that might produce some of the same effects as CR even if it is not quite as beneficial as the real thing.
One way to achieve the beneficial effects of CR without the muscle wasting, decreased mobility, and osteoporosis side effects is to use a caloric restriction mimetic. The best-known ones so far have been low dose 2-Dexoyglucose (2DG), resveratrol, rapamycin, and PQQ.
1. 2-Deoxyglucose (2DG) - A diet of 0.3%-0.4% 2DG does not affect body weight or food intake, but induces many of the biochemical and physiological effects of CR, such as lowered insulin levels, reduced body temperature, and lowered rates of tumor development.
2. Resveratrol - A 30 day, prospective, randomized, placebo controlled study of obese men given 150mg of Reseveratrol per day showed many of the benefits of CR. Resveratrol reduced sleeping and and resting metabolic rate. In muscle, Resveratrol increased AMPK, SIRT1, and PGC-1a levels. It also improved muscle respiration on mitochondrial fatty acid metabolism. Reseveratrol decreased circulating glucose, triglycerides, alanine-aminotransferase, and inflammation markers. Systolic BP dropped and the HOMA index improved.
At the xenohormesis bar, you eat plants and substances from plants that are powerful defensive molecules developed over millions of years in the plants to protect themselves against stresses. These substances activate evolutionarily-conserved pathways in you, so the plant’s defensive mechanisms become part of your own when you eat them. See the PART 1: Slaying Two Dragons with One Stone blog entry, the section on the Concept of Xenohormesis.
This bar is based on the principle that if I eat a powerful enemy of my enemy, he will become part of me and help me defeat my enemy. And it is based on the assumption that this will still work even if the enemy of my enemy is of a very different species, a plant in this case. Anoush was a powerful Persian warrior who had known only victory through battles.
Anoush loved his mother Atosa dearly and when she became very sickly he asked “What is the evil spirit that has occupied my mother, the most-beautiful Atosa? Tell me so I can defeat it, be it by spear, dagger or sword.” He had no answer until one day a wizened old medicine man told him “Your mother has internalized the spirit of Carcinogotcha. Your sword and dagger are useless to defeat Carcinogotcha. But your answer lies in the Turmeric plant. A powerful warrior Oncogone was banished in most ancient times into the roots of that plant because he insulted the supreme god Ahura Mazda. The roots of the Turmeric plant now possess the spirit of Oncogone, and this spirit can defeat your mother’s Carcinogocha,
Feed Atosa amply of the root of Turmeric, and Oncogone will defeat Carninogotcha and she will be well.” Anoush dug up Turmeric roots with his sword and did exactly this. Then his mother Atosa rapidly improved and started to go to Zumba classes. So here was born the idea of Xenohormesis, a process through which a plant’s health stress defenses can become part of the stress defenses of those who partake of the plant.
These menu items are for those who would rather eat than exercise, people who would never patronize the Oxygen Bankruptcy Bar. Although our philosophy is that there is no “true exercise mimetic”, these natural compounds probably do the most to give the organism the ability to “have their cake and eat it too.” The drinks at the Xenohormetic bar range from pleasant-tasting ones to bitter ones. The compounds include most of the health-inducing phytosubstances written about in previous blog entries: bitter melon, gamboic acid, ginger, curcumin, resveratrol, green tea, extra-virgin olive oil secoiridoids, sulphorane from broccoli, etc. And a bunch of lesser-known ones are offered as specials. This week, featured drinks are based on exotic mushrooms.
Pathway to be Altered – Example phytochemical or class of natural compounds, Biomarker
1. Nrf2 pathway activation – isothiocyanates, breathalyzer
16. AP-1 inhibition – plumbagin, curcumin, isothiocyanates , Not sure of best ones
17. JAK and STAT3 inhibition – curcurbitacin, indirubin, plumbagin, deoxytetrangomycin IL-6, Not sure of best ones
18. Beta-catenin inhibition – fisetin, Not sure of best ones
19. c-Myc inhibition – we don’t know of any yet
20. Angiogenesis inhibitors (VEGF) – fish oil, krill oil, emodin, coleon A lactone, gentisic acid
III. The Xenobiotic Bar
A xenobiotic is a substance which is a stranger in an organism into which it is introduced; it is not normally produced or expected to be or present there and is not part of a normal diet. Most drugs are xenobiotics. So are many food additives.
This part of the Hormesis Bar will probably be the most popular, since it is the only area in the bar that serves alcohol. Although the alcohol is our most popular menu item, we have other items on the menu that are not for the “faint of heart”. Rather, this is only for “true believers” who believe that “pinches of poison” are actually good for you. These “poisons” have been shown to have a positive effect on health at low, hormetic doses and include carbon monoxide, arsenic, and even emotional stress doses administered by a simulated judgemental father-in-law who keeps telling you what a loser you are:
A. Alcohol - alcohol is the most well studied poison and the most well-used poison in all cultures. In 2012, UCLA researchers showed that 0.005% of ethanol doubled the lifespan of C. elegans. This is the equivalent of 1mM blood alcohol levels. Doses of higher than 0.4% provided no longevity benefit. Since the legal limit for driving is 0.08 or 8%, this means that you can drive your car with the hormetic dose of alcohol on board. This means that we need no designated drivers at the Xenobiotic bar. In humans, low dose alcohol has shown to have a protective effect against heart disease, stroke, and possibly some types of cancer. The “low dose” is probably less than 1/2 glasses per day. No one will get drunk or pregnant at this dose. The mechanisms by which this is beneficial are thought to be largely due to adaptive responses to the stress of alcohol (i.e. hormetic effect)
B. UV light - UV light has been demonized to be the cause of skin cancer. To the contrary, low doses of UV light have been shown to have several benefits. The most well known is the positive effects of UV light on the synthesis of Vitamin D. What has not been publicized well is the recent study in 2005 that showed that low doses of sunlight can actually reduce the risk of skin cancer. We intend to “serve UV light” up at the Xenobiotic Bar, but in controlled doses for limited windows of time.
C. Methyl mercury -This was a popular menu item 2,300 years ago when Emperor Quin was in power in China). In fact mercury has gone out of style since the Chinese emperors all died off. Nevertheless, a 2010 study in Mallard ducks showed that low doses of Methyl mercury actually improved the hatching rate of Mallard eggs. You can get it through heaping portions of tuna fish and swordfish.
D. Dioxin - The compound that is related to the Vietnam era poison, Agent Orange, is one of the most vilified toxins in the field of toxicology. In rats, low dose dioxin has actually been shown to reduce the incidence of tumors, rather than increasing the incidence of cancer. This one requires signing a special release form.
E. Allergy antigen exposure - Most people try to avoid hay fever allergens. At the Xenobiotic Bar, small doses of hay fever allergens are constantly being blown all over the bar stools and counters and onto patrons there. There is good scientific evidence that “Allergen immunotherapy”, which is the practice of introducing low doses of an allergen to a patient and gradually increasing the dose to build up tolerance. This form of allergy medicine is practiced clinically today with good results. According to Dr. Linda Cox, chair of the Immunotherapy and Diagnostics Committee of the American College of Allergy, Asthma and Immunology (ACAAI), studies in Germany and the U.S. showed significantly improved cost effectiveness of allergen immunotherapy compared with conventional anti-allergy drugs in treating allergic rhinitis and asthma.
F. ROS- Contrary to what most people are trying to do, we here at the Xenobiotic Bar believe that free radicals are not really “xenobiotic compounds”, but since they are viewed as “toxins” by much of the scientific establishment, we intend to put “ROS” on our list of xenobiotic compounds. However, instead of trying to “get rid of ROS”, we “promote ROS” as a positive signaling molecule with wonderful health benefits. We intend to serve hydrogen peroxide (H2O2) as a “smokeless aerosolized cigarette,” a “peroxide on the rocks” drink, and even a H2O2 intravenous drip. The key is “getting the dose right”.
CONCLUSION
We believe that it is time to “practice what we preach.”
We believe that there are three categories of “low dose stressors” that should be employed in any program that is designed to promote health span, reduce cancer, and increase longevity. The three categories are Hormesis, Xenohormesis, and Xenobiotics. We believe that all of these pathways should be exploited and activated via appropriate doses. Although “stress mimetics” such as xenohormetic food ingredients or xenobiotic toxins are good, they are not a true substitute for the “real stressor”. That is why we intend to serve up heat, cold, hypoxia, exercise, starvation, and radiation at the hormesis bar. The xenohormesis bar will also be included to cover as many pathways as is feasible.
WARNING
Again, do not try these “drinks” or procedures for yourself unless you fully know what you are doing. And we take no responsibility for any consequences if you do.
MEDICAL DISCLAIMER
FROM TIME TO TIME, THIS BLOG DISCUSSES DISEASE PROCESSES. THE INTENTION OF THOSE DISCUSSIONS IS TO CONVEY CURRENT RESEARCH FINDINGS AND OPINIONS, NOT TO GIVE MEDICAL ADVICE. THE INFORMATION IN POSTS IN THIS BLOG IS NOT A SUBSTITUTE FOR A LICENSED PHYSICIAN’S MEDICAL ADVICE. IF ANY ADVICE, OPINIONS, OR INSTRUCTIONS HEREIN CONFLICT WITH THAT OF A TREATING LICENSED PHYSICIAN, DEFER TO THE OPINION OF THE PHYSICIAN. THIS INFORMATION IS INTENDED FOR PEOPLE IN GOOD HEALTH. IT IS THE READER’S RESPONSIBILITY TO KNOW HIS OR HER MEDICAL HISTORY AND ENSURE THAT ACTIONS OR SUPPLEMENTS HE OR SHE TAKES DO NOT CREATE AN ADVERSE REACTION
Quorum sensing (QS) is a fundamental biological process involving how cells in assemblages sense and communicate and cooperate with each other. It applies to bacterial colonies as well as to animal cells in organs. This Part 1 blog entry relates to survival-based signaling among bacteria. It is possible that the ravages of many infectious diseases can be averted by the use of substances to interrupt QS signaling. Plant-based substances hold particular promise in this regard. This blog covers current and recent research related to this hypothesis. Much of the relevant research is quite new and I cite several 2013 publications. A Quorum Sensing Part 2 blog entry will be concerned with important roles of QS in animal and human cells including how deregulated QS may lead to tumorigenesis and the important roles of QS in stem cell maintenance and differentiation.
Along with the discovery of quorum sensing came the realization that infectious bacteria are highly social entities who cooperate in multiple ways for their survival. The 2013 publication Impacts of quorum sensing on microbial metabolism and human health provides an overview: “Bacteria were considered to be lonely ‘mutes’ for hundreds of years. However, recently it was found that bacteria usually coordinate their behaviors at the population level by producing (speaking), sensing (listening), and responding to small signal molecules. This so-called quorum sensing (QS) regulation enables bacteria to live in a ‘society’ with cell-cell communication and controls many important bacterial behaviors. In this chapter, QS systems and their signal molecules for Gram-negative and Gram-positive bacteria are introduced. Most interestingly, QS regulates the important bacterial behaviors such as metabolism and pathogenesis. QS-regulated microbial metabolism includes antibiotic synthesis, pollutant biodegradation, and bioenergy production, which are very relevant to human health. QS is also well-known for its involvement in bacterial pathogenesis, such as iin nfections by Pseudomonas aeruginosa and Staphylococcus aureus. Novel disease diagnosis strategies and antimicrobial agents have also been developed based on QS regulation on bacterial infections. In addition, to meet the requirements for the detection/quantification of QS signaling molecules for research and application, different biosensors have been constructed, which will also be reviewed here. QS regulation is essential to bacterial survival and important to human health. A better understanding of QS could lead better control/manipulation of bacteria, thus making them more helpful to people.”
What is quorum sensing? The basics related to bacteria
Quorum sensing (QS) is a signaling process used by many Gram-negative and Gram-positive bacteria to monitor their own population density and coordinate gene regulation so as to determine collective behavior. By knowing how many other bacterial cells are present, bacteria can trigger genes that radically affect individual cell behavior and as a result behavior of a whole colony of cells. For example a colony of bioluminescent bacteria can remain dark until quorum sensing determines there is a critical density of such bacteria, at which point the entire colony lights up at once. Another example of action that can result from a quorum sensing trigger point is cells starting to secrete substances that will make up a protective biofilm for a cell colony. Another cell action that can be triggered by quorum sensing in many bacteria is the release of cytotoxic chemicals that attack a host organism. Based on the quorum sensing, essentially all the bacteria switch at once to being aggressive warriors.. This strategy is used, for example by Pseudomonas aeruginosa, which causes infections in patients with compromised immune systems and cystic fibrosis. The bacteria do not launch an attack on a patient until their quorum sensing mechanisms tell them that they are in sufficient numbers to possibly succeed. Then they attack all at once. The evolutionary advantage to the bacteria is that they can hide out without being noticed in the host organism until they have multiplied sufficiently. If they don’t maintain a low profile, they could be wiped out in the initial stage by the hosts immune system. These are natural “sleeper cells.”
Signaling is via cell-secreted signaling molecules called autoinducers. Each autoinducer signaling molecule is highly specific to the bacterial species involved, and activates cell-membrane sensors in other bacteria of the same species. In the case of Gram-negative bacteria N-acyl homoserine lactone (AHL) autoinducer signal molecules are employed. Quorum-sensing circuits have been identified in over 25 species of Gram-negative bacteria. “Gram-positive bacteria employ secreted peptides processed from precursors, that can be used as autoinducers for QS — Signals are actively exported outside the cell, where they interact with the external domains of membrane-bound sensor proteins. The transduction of a signal generated by a phosphorylation cascade culminates in the activation of a DNA-binding protein, that influences the transcription of specific genes, so that each sensor protein is highly selective for a given peptide signal. In Gram-positive bacteria, a peptide signal precursor locus is translated into a precursor protein that is cleaved to produce the processed peptide autoinducer signal. This signal is then usually transported out of the cell via an ABC transporter(from Quorum Sensing and Phytochemicals, 2013).”
Quorum sensing is now recognized as a critical phenomenon in many infectious disease processes and Pubmed.org now lists 4194 research publications on QS going back to 1994. A lively and easily understandable video describing QS in bacteria can be found here.
There are additional kinds of quorum sensing and functions for it. An example is allowing members of a bacterial species to sense not only how many of its own kind are in the environment but also the total number of bacteria of any kind in the environment. This can allow the cell to feel out what the competitive environment is like and help it in deciding whether and when to trigger gene activation actions. Moreover, actions can go beyond biofilm formation or releasing biotoxic factors. The publication Quorum sensing and disease reports: “Much has been learned in the past decade about the complex behavior of bacteria under the influence of various Quorum Sensing systems. Many bacteria possess multiple QS systems, which control different aspects of virulence or other behaviors at different times in the infection process. And each bacteria species responds differently to QS stimuli, depending on environmental pressures, the presence or absence of competing bacterial species, the immune response of the host and other conditions. – For example, many bacteria form dense collections of cells called biofilms that are enclosed in a protective barrier. Such biofilms are highly resistant to host immune responses, anti-bacterial agents and other threats. The creation of these biofilms is regulated by QS signaling, and they generally are created after very large numbers of bacteria are present. – Some bacteria, however, use quorum sensing to end the biofilm stage as well as begin it. Cholera is one such example. In the final stages of infection, cholera uses QS signaling to dissolve existing biofilms and begin a massive dispersal of individual bacteria. This creates a highly infectious condition within the host and aids in cholera’s ability to spread. – There are many such examples of different bacteria responding to QS signaling in diverse ways at different times during the infection process. – What seems to be nearly universal, however, is the bacteria’s tendency to maintain as low a profile as possible when the host is first infected. It is believed that this behavior minimizes the host’s immune response while the bacteria build sufficient numbers to mount an effective offense. When a sufficient number is reached, the bacteria manifest their pathogenic (disease causing) arsenal. The timing of this transition reveals an exquisite genetic intelligence: when pathogenic bacteria have reached a density that would trigger the host’s full-scale immune response whether or not there existed a disease symptom stimulus, the bacteria, using QS signaling, begin their disease-causing phase. The disease stresses the host, thereby weakening the host’s ability to respond to the infection. This gives advantage to the bacteria.”
Among the actions resulting from quorum sensing signaling, certain bacteria can generate surfactants and extra flagella which facilitate bacterial “swarming” across solid surfaces(ref)(ref).
A diversity of related quorum sensing populations can exist even for a single pathogen.
The 2009 e-publication Instantaneous within-patient diversity of Pseudomonas aeruginosa quorum-sensing populations from cystic fibrosis lung infectionsreported: “To obtain insight into the instantaneous within-patient diversity of QS, we assayed a panel of 135 concurrent P. aeruginosa isolates from eight different adult CF patients (9 to 20 isolates per patient) for various QS-controlled phenotypes. Most patients contained complex mixtures of QS-proficient and -deficient isolates. Among all patients, deficiency in individual phenotypes ranged from 0 to about 90%. Acyl-HSL, sequencing, and complementation analyses of variants with global loss-of-function phenotypes revealed dependency upon the central QS circuitry genes lasR, lasI, and rhlI. Deficient and proficient isolates were clonally related, implying evolution from a common ancestor in vivo. Our results show that the diversity of QS types is high within and among patients, suggesting diverse selection pressures in the CF lung. A single selective mechanism, be it of a social or nonsocial nature, is unlikely to account for such heterogeneity. The observed diversity also shows that conclusions about the properties of P. aeruginosa QS populations in individual CF infections cannot be drawn from the characterization of one or a few selected isolates.”
There is much more to be said about quorum sensing in bacteria as will be unfolding in the course of this and the Part 2 blog entries.
To maintain perspective, we need to keep in mind that most microbes humans encounter are either positive or neutral for our health, and that quorum sensing is an evolutionarily-conserved property of many families of microbes, including all colony-forming ones.
Much of the research and literature related to quorum sensing has focused on pathological bacteria and how inhibiting quorum sensing might be a good strategy for averting or curing diseases. However, dealing with infectious diseases is only part of what must be taken into account to ensure health and longevity. Maintaining quorum sensing in beneficial bacteria is important too.
It is estimated that there are approximately a10 times more foreign cells in the human body than our actual human cells, some 100 trillion of them compared to 10 trillion of our own. These foreign cells are of thousands of species and exist in multiple biomes including the gut biome, the skin biome, the mouth biome, the vaginal biome, etc. I discussed just the gut biome in the blog entry Gut microbiota, probiotics, prebiotics and synbiotics – keys to health and longevity. A human typically carries thousands of species of such microbes, and the species can vary significantly depending on the local biome concerned. For example, the skin biomes for the left-hand and for the right hand may mostly consist of different bacteria. The “not us” microbes fall generally into three categories: 1. ones that exist in harmony with us that are known to play positive roles in health maintenance, 2. bacteria that generally play a positive or neutral role with respect to our health but which can turn to play negative roles(ref). Such bacteria are called commensalbacteria, where the bacteria depends on a symbiotic relationship with the host with little or no harm to the host. And 3. bacteria associated with pathologies, ones like Salmonella, Listeria, Pseudomonas, Staphylococcus and Lactobacillus. Traditional medical research has focused mainly on bacteria in category 3. The study of human biomes did not get really underway until the late 1990s, and the roles these biomes play with regard to our health are only now being explored.
In general, animal biomes and the bacteria in them have co-evolved. Bacteria have generally worked out a friendly or neutral symbiotic state with the host organism and with each other. Bacteria in the human biomes contribute in multiple ways to both our health and longevity. Many of these bacteria employ quorum sensing as part of their basic survival approach, certainly the ones that are colony-forming. Human cells and multiple bacteria may exist in the same organ, either cooperating with each other, indifferent or competing, and all using their own mechanisms of quorum sensing. In this blog entry, consistent with emphasis in the literature, we concentrate on the relatively small number of bacteria which are pathological, and focus on mechanisms for controlling or curing diseases via inhibiting quorum sensing. However, the last thing we want to do is inhibit quorum sensing across-the-board. Doing that would stop the actions of multiple beneficial microbes as well as essential QS among our own cells and probably be disastrous for health.
Image source Bacterial species involved with dental biofilm accumulation and tooth decay. All employ quorum sensing.
The mechanisms and activities of QS are being actively researched. QS is a hot topic.
For example, the June 2013 publication Novel quorum-sensing peptides mediating interspecies bacterial cell death reports: “Escherichia coli mazEF is a toxin-antitoxin stress-induced module mediating cell death. It requires the quorum-sensing signal (QS) “extracellular death factor” (EDF), the penta-peptide NNWNN (EcEDF), enhancing the endoribonucleolytic activity of E. coli toxin MazF. Here we discovered that E. coli mazEF-mediated cell death could be triggered by QS peptides from the supernatants (SN) of the Gram-positive bacterium Bacillus subtilis and the Gram-negative bacterium Pseudomonas aeruginosa. In the SN of B. subtilis, we found one EDF, the hexapeptide RGQQNE, called BsEDF. In the SN of P. aeruginosa, we found three EDFs: the nonapeptide INEQTVVTK, called PaEDF-1, and two hexadecapeptides, VEVSDDGSGGNTSLSQ, called PaEDF-2, and APKLSDGAAAGYVTKA, called PaEDF-3. When added to a diluted E. coli cultures, each of these peptides acted as an interspecies EDF that triggered mazEF-mediated death. Furthermore, though their sequences are very different, each of these EDFs amplified the endoribonucleolytic activity of E. coli MazF, probably by interacting with different sites on E. coli MazF. Finally, we suggest that EDFs may become the basis for a new class of antibiotics that trigger death from outside the bacterial cells. IMPORTANCE Bacteria communicate with one another via quorum-sensing signal (QS) molecules. QS provides a mechanism for bacteria to monitor each other’s presence and to modulate gene expression in response to population density. Previously, we added E. coli EDF (EcEDF), the peptide NNWNN, to this list of QS molecules. Here we extended the group of QS peptides to several additional different peptides. The new EDFs are produced by two other bacteria, Bacillus subtilis and Pseudomonas aeruginosa. Thus, in this study we established a “new family of EDFs.” This family provides the first example of quorum-sensing molecules participating in interspecies bacterial cell death. Furthermore, each of these peptides provides the basis of a new class of antibiotics triggering death by acting from outside the cell.”
A favorite bacterium for study of QS dynamics has been Chromobacterium violaceum –“a Gram-negative, facultative anaerobic, non-sporing coccobacillus. It is part of the normal flora of water and soil of tropical and sub-tropical regions of the world. It produces a natural antibiotic called violacein, which may be useful for the treatment of colon and other cancers.[1] It grows readily on nutrient agar, producing distinctive smooth low convex colonies with a dark violet metallic sheen (due to violacein production). Its full genome was published in 2003.[2] It has the ability to break down tarballs.[3] (ref)”(ref). For example, Chromobacterium violaceum CV026 is sometimes used as a reporter strain for measuring QS inhibitory activity of candidate substances.
Inhibiting quorum sensing could be a fast and effective way of stopping and clearing up an infectious disease. The need is very significant.
Starting more than10 years ago, there has been an increasing interest in averting or curing infectious diseases through inhibition of quorum sensing. One reason for this has been increasing resistance of strains of pathogenic bacteria to conventional antibiotics, and an inability of the pharmaceutical industry to develop conventional-antibiotics to replace ones that are no longer effective against bacteria which have evolved to becoming drug-resistant. Antibiotic resistance is a growing crises(ref). Another reason is that interfering with pathogen signaling seems to be novel, potentially both safe and effective, and a possible way to outwit bacteria who are smart at evolving.
The March 2013 publication Quorum sensing inhibitors: an overview provides a summary: “Excessive and indiscriminate use of antibiotics to treat bacterial infections has lead to the emergence of multiple drug resistant strains. Most infectious diseases are caused by bacteria which proliferate within quorum sensing (QS) mediated biofilms. Efforts to disrupt biofilms have enabled the identification of bioactive molecules produced by prokaryotes and eukaryotes. These molecules act primarily by quenching the QS system. The phenomenon is also termed as quorum quenching (QQ). In addition, synthetic compounds have also been found to be effective in QQ. This review focuses primarily on natural and synthetic quorum sensing inhibitors (QSIs) with the potential for treating bacterial infections. It has been opined that the most versatile prokaryotes to produce QSI are likely to be those, which are generally regarded as safe. Among the eukaryotes, certain legumes and traditional medicinal plants are likely to act as QSIs. Such findings are likely to lead to efficient treatments with much lower doses of drugs especially antibiotics than required at present.”
Another attractive aspect of QS inhibition as a potential strategy against infectious diseases appeared to be how quickly and effectively it could work, potentially with no side effects. Again, from Quorum sensing and disease: “The second, nearly universal trait, is that even after a bacterium has entered its pathogenic phase, it quickly returns to its non-pathogenic state when we disable its QS mechanisms. This is another clear sign of genetic intelligence, for when a bacterium in its pathogenic state is passed on to a new host, the new host would quickly locate and dispatch the invader if the invader continued in its pathogenic form. What happens when such transfers do occur is that, from the bacterium’s point of view, there is an immediate cessation of the quorum. This is the indication to the bacterium that it is in a new host, and that the infection process is starting over. All pathogenic activities of the bacterium then cease, giving the bacterium time to multiply and, later, mount another offense. This is the beauty of the anti-QS approach to the treatment of infection. It terminates the disease causing activities, thereby returning the body to strength and health. The body can then deal with the infection through its normal immune system responses, without inflammation or other deleterious effects. In contrast, current antibiotic treatments attempt to kill the bacteria while the body is in a state that is already compromised by the disease. The combination of the disease and the stress caused by many antibiotics can further weaken the system to dangerous levels.”
But the road was discovered to be bumpy. The 2004 document Quorum-sensing control in Staphylococci – a target for antimicrobial drug therapy? expresses this perception and warns of a case where QS inhibition did not appear to work: “Today, we find ourselves in an urgent need for novel antibacterial drugs, as many important human pathogens have acquired multiple antibiotic resistance factors. Among those, Staphylococcus aureus and S. epidermidis play a major role as the leading sources of nosocomial infections. Recently, it has been suggested to develop therapeutics that attack bacterial virulence rather than kill bacteria. Such drugs are called “antipathogenic” and are believed to reduce the development of antibiotic resistance. Specifically, cell-density-dependent gene regulation (quorum-sensing) in bacteria has been proposed as a potential target. While promising reports exist about quorum-sensing blockers in gram-negative bacteria, the use of the staphylococcal quorum-sensing system as a drug target is now seen in an increasingly critical way. Inhibition of quorum-sensing in Staphylococcus has been shown to enhance biofilm formation. Furthermore, down-regulation or mutation of the Staphylococcus quorum-sensing system increases bacterial persistence in device-related infection, suggesting that interference with quorum-sensing would enhance rather than suppress this important type of staphylococcal disease. The chemical nature and biological function of another proposed staphylococcal quorum-sensing inhibitor, named “RIP”, are insufficiently characterized. Targeting quorum-sensing systems might in principle constitute a reasonable way to find novel antibacterial drugs. However, as outlined here, this approach requires careful investigation in every specific pathogen and type of infection.”
It appears that a number of phytosubstances are effective at inhibiting QS.
It has been known for a long time that a number of phyto substances (plant-based substances) have been effective in combating a number of infectious diseases(ref)(ref). And this wisdom is basic in traditions of folk and herbal medicine. However, the mechanisms used by phytochemicals to do this have not been clear. Research during the last ten years indicates that many of these phytochemicals effectively inhibit quorum sensing in pathogenic target bacteria, as will be elaborated below. This hitherto unidentified mechanism could possibly become very important for dealing with infectious diseases. We have written in this blog extensively about the potential roles of phyto substances in averting cancer and aging(ref)(ref)(ref)(ref), epigenetic pathways through which phyto substances work(ref)(ref), and the potential role they could play in treating age-related neurological diseases(ref). This is the first blog entry, however, seriously relating phyto substances to infectious diseases.
The fact that phytochemicals, substances developed in plants as stress defenses, can also work in humans to deal with similar stresses via inhibition of QS is another example of Xenohormesis. See the discussion in the Xenohormesis section of this blog entry.
Starting around ten years ago, researchers have been looking into various medicinal plants and phytochemicals to determine whether they employ QS as an antimicrobial strategy. They have identified a rather large number of them. It appears that QS inhibition of pathogenic bacteria is an important survival strategy used by many plants.
The 2007 publication Dietary phytochemicals as quorum sensing inhibitors reported: “Quorum sensing (QS) is a cell density dependent expression of species in bacteria mediated by hormone like compounds called autoinducers (AI). Several processes responsible for successful establishment of bacterial infection are mediated by QS. Inhibition of QS is therefore being considered as a new target for antimicrobial chemotherapy. Dietary phytochemicals are secondary metabolites in plants known to have several health benefits including antimicrobial activity. However, their ability to inhibit QS has never been studied. Our objective was to investigate the effect of sub-lethal concentrations (SLC) of bioactive dietary phytochemical extracts from common dietary fruit, herb and spice extracts on modulating QS mediated by AI in model bioassay test systems. QS inhibition was measured in violacein pigment producing Chromobacterium violaceum O26 (CVO26) and CV 31532 system, mediated by AI known as acylated homeserine lactone (AHL). We also investigated the effect of the sub-lethal concentrations of the extracts on swarming motility of pathogens Escherichia coli (EC)O157:H7 and Pseudomonas aeruginosa (PA-01). Our results indicate that all extracts significantly inhibited quorum sensing. The mechanism of inhibition appeared to be combination of interfering with AHL activity and modulating the synthesis of AHL’s. Our results also indicated that various phytochemical extracts which inhibited QS also inhibited swarming of pathogenic bacteria, known to be modulated by QS. The observation that phytochemicals from foods can inhibit QS related processes opens up an exciting new strategy for antimicrobial chemotherapy and lead to the discovery of new category of antibiotics which can overcome the issues related to antimicrobial resistance.”
It makes sense. Because QS is a common evolutionary-driven strategy employed by pathogenic bacteria, plants in turn have evolved a broad repertory of phytosubstances which inhibit QS.
The May 2013 review Plant-derived natural products as sources of anti-quorum sensing compounds explains this point: “Just like animals and humans, plants are constantly exposed to bacterial infections, it is therefore logical to expect that plants have developed sophisticated chemical mechanisms to combat pathogens. In this review, we have surveyed the various types of plant-based natural products that exhibit anti-quorum sensing properties and their anti-quorum sensing mechanisms.”
The 2013 publication Quorum Sensing and Phytochemicals reviews mechanisms of QS employed by both gram-negative and gram-positive bacteria and discusses another important topic: the potential role of QS inhibition to slow or prevent food spoilage due to bacteria. “Most infectious diseases are caused by bacteria, which proliferate within quorum sensing (QS)-mediated biofilms. Efforts to block QS in bacteria and disrupt biofilms have enabled the identification of bioactive molecules that are also produced by plants. This mini review primarily focuses on natural QS inhibitors, which display potential for treating bacterial infections and also enhance the safety of food supply.” A great many specific phytochemicals are mentioned in the review.
A sample of recent research publications descriptive of or related to QS inhibition by phytosubstances is:
Marine organisms like sponges might also be good sources of QS inhibitors that would work against human pathogens. The 2012 publication Inhibition of Quorum Sensing Mediated Virulence Factors Production in Urinary Pathogen Serratia marcescens PS1 by Marine Spongesreports: “The focal intent of this study was to find out an alternative strategy for the antibiotic usage against bacterial infections. The quorum sensing inhibitory (QSI) activity of marine sponges collected from Palk Bay, India was evaluated against acyl homoserine lactone (AHL) mediated violacein production in Chromobacterium violaceum (ATCC 12472), CV026 and virulence gene expressions in clinical isolate Serratia marcescens PS1. Out of 29 marine sponges tested, the methanol extracts of Aphrocallistes bocagei (TS 8), Haliclona (Gellius) megastoma (TS 25) and Clathria atrasanguinea (TS 27) inhibited the AHL mediated violacein production in C. violaceum (ATCC 12472) and CV026. Further, these sponge extracts inhibited the AHL dependent prodigiosin pigment, virulence enzymes such as protease, hemolysin production and biofilm formation in S. marcescens PS1. However, these sponge extracts were not inhibitory to bacterial growth, which reveals the fact that the QSI activity of these extracts was not related to static or killing effects on bacteria. Based on the obtained results, it is envisaged that the marine sponges could pave the way to prevent quorum sensing (QS) mediated bacterial infections.”
Some conventional drugs also inhibit QS in pathogenic bacteria.
The antibiotic doxycycline appears to have interference with QS as one of its mechanisms of action. The publication Doxycycline interferes with quorum sensing-mediated virulence factors and biofilm formation in Gram-negative bacteria reports: “Inhibition of quorum sensing (QS)-regulated virulence factors including biofilm is a recognized anti-pathogenic drug target. The search for safe and effective anti-QS agents is expected to be useful to combat diseases caused by multidrug-resistant bacteria. In this study, effect of a commonly used antibiotic, doxycycline on QS was evaluated using sensor strains of Chromobacterium violaceum (ATCC 12472 and CVO26) and Pseudomonas aeruginosa PAO1. Sub-MICs of doxycycline reduced QS-controlled violacein production in C. violaceum to a significant degree (70 %) and showed a significant reduction of LasB elastase (67.2 %), pyocyanin (69.1 %), chitinase (69.8 %) and protease (65 %) production and swarming motility (74 %) in P. aeruginosa PAO1 over untreated controls. Similar results were also recorded against a clinical strain of P. aeruginosa (PAF-79). Interestingly, doxycycline at respective sub-MICs (4 and 32 μg ml(-1)) significantly reduced the biofilm-forming capability and exopolysaccharide production in both the strains of P. aeruginosa (PAO1 and PAF-79) over untreated controls. The results of this study highlight the multiple actions of doxycycline against QS-linked traits/virulence factors and its potential to attenuate virulence of P. aeruginosa.”
Also you can see New life for an old drug: the anthelmintic drug niclosamide inhibits Pseudomonas aeruginosa quorum sensing. “Microarray analysis showed that niclosamide affects the transcription of about 250 genes, with a high degree of target specificity toward the QS-dependent regulon. Phenotypic assays demonstrated that niclosamide suppresses surface motility and production of the secreted virulence factors elastase, pyocyanin, and rhamnolipids, and it reduces biofilm formation. In accordance with the strong antivirulence activity disclosed in vitro, niclosamide prevented P. aeruginosa pathogenicity in an insect model of acute infection.”
Some antibiotics, acting as signaling molecules, can in a concentration-dependent manner actually promote QS in bacteria
The April 2013 publication Antibiotics at subinhibitory concentrations improve the quorum sensing behavior of Chromobacterium violaceumrelates: “Increasing evidence has shown that antibiotics function as intermicrobial signaling molecules instead of killing weapons. However, mechanisms and key factors that are involved in such functions remain poorly understood. Earlier findings have associated antibiotic signaling with quorum sensing (QS); however, results varied among experiments, antibiotics, and bacterial strains. In this study, we found that antibiotics at subinhibitory concentrations improved the violacein-producing ability of Chromobacterium violaceum ATCC 12472. Quantitative real-time polymerase chain reaction of QS-associated gene transcripts and bioassay of violacein production in a QS mutant strain demonstrated that antibiotics enhanced the production of N-acyl-L-homoserine lactones (AHLs; QS signaling molecules) and increased AHL-inducing QS-mediated virulence, including chitinase production and biofilm formation. Moreover, a positive flagellar activity and an increased bacterial clustering ability were found, which are related to the antibiotic-induced biofilm formation. Our findings suggested that antibiotic-mediated interspecific signaling also occurs in C. violaceum, thereby expanding the knowledge and language of cell-to-cell communication.” The implication is that some antibiotics may exercise their influence via a hormetic biphasic dose-response curve, killing bacteria when the dose is super-hormetic and acting as signaling molecules when the dose is in the low hormetic range.
There is also work on synthesizing compounds for inhibiting QS.
For example, the March 2013 publication Design, synthesis and biological evaluation of N-sulfonyl homoserine lactone derivatives as inhibitors of quorum sensing in Chromobacterium violaceum reports “A novel series of N-sulfonyl homoserine lactone derivatives 5a-l has been designed, synthesized and evaluated for quorum sensing inhibitory activities towards violacein production. Of the compounds synthesized, compound 5h was found to possess an excellent level of enantiopurity (99.2% e.e.). The results indicated that compounds bearing an ortho substituent on their phenyl ring exhibited excellent levels of inhibitory activity against violacein production. Compounds 5h and 5k in particular, with IC₅₀ values of 1.64 and 1.66 µM, respectively, were identified as promising lead compounds for further structural modification.”
There are other important applications of biofilm inhibition by inhibiting QS using natural substances
Inhibition of QS has uses beyond those addressing human bacterial pathologies. I have already mentioned the food-preservation application. There are also marine applications. An example is keeping boat bottoms and propellers free of fouling by colonies of marine bacteria, For example, the June 2013 publication Coral-associated bacteria, quorum sensing disrupters, and the regulation of biofouling reports: “Marine biofouling, the settlement of microorganisms and macroorganisms on structures submerged in seawater, although economically detrimental, is a successful strategy for survival in hostile environments, where coordinated bacterial communities establish biofilms via the regulation of quorum sensing (QS) communication systems. The inhibition of QS activity among bacteria isolated from different coral species was investigated to gain further insight into its potency in the attenuation, or even the prevention, of undesirable biofouling on marine organisms. It is hypothesized that coral mucus/microorganism interactions are competitive, suggesting that the dominant communities secrete QS disruptive compounds. One hundred and twenty bacterial isolates were collected from healthy coral species and screened for their ability to inhibit QS using three bioreporter strains. Approximately 12, 11, and 24% of the isolates exhibited anti-QS activity against Escherichia coli pSB1075, Chromobacterium violaceum CV026, and Agrobacterium tumefaciens KYC55 indicator strains, respectively. Isolates with positive activity against the bioluminescent monitor strains were scanned via a cytotoxic/genotoxic, E. coli TV1061 and DPD2794 antimicrobial panel. Isolates detected by C. violaceum CV026 and A. tumefaciens KYC55 reporter strains were tested for their ability to inhibit the growth of these reporter strains, which were found to be unaffected. Tests of the Favia sp. coral isolate Fav 2-50-7 (>98% similarity to Vibrio harveyi) for its ability to attenuate the formation of biofilm showed extensive inhibitory activity against biofilms of Pseudomonas aeruginosa and Acinetobacter baumannii. To ascertain the stability and general structure of the active compound, cell-free culture supernatants exposed to an increasing temperature gradient or to digestion by proteinase K, were shown to maintain potent QS attenuation and the ability to inhibit the growth of biofilms. Mass spectrometry confirmed the presence of a low molecular mass compound. The anti-QS strategy exemplified in the coral mucus is a model with potentially wide applications, including countering the ecological threat posed by biofilms. Manipulating synchronized bacterial behavior by detecting new QS inhibitors will facilitate the discovery of new antifouling compounds.”
Although most discussions of QS in bacteria are framed in terms of free-floating signal molecules activating cell surface receptors, there is likely to be more to the picture. These signaling molecules may be in some or most instances packaged into vesicles which play a similar role to that played by exosomes in animal species.
From a 2008 news release in The Times Microbial Membrane Vesicles in Bacteria: Taking Quorum Sensing in New Directions: “Commonly, quorum sensing molecules are lactones, small peptides, or small lipids. It is thought that these small molecules are secreted by the cell and diffuse through the aqueous environment where they can interact with other cells. This is all well and good, IF the molecule of interest is hydrophillic and freely diffusible. But researchers are finding that many quorum sensing molecules are NOT hydrophillic, and instead very hydrophobic, such as a quinilone with a long chain fatty acid attached. One such example of this, is the Pseudomonas aeruginosa molecule called PQS. — How can a molecule be used as an extracellular signal, if it can not diffuse freely? Dr. Marvin Whitely and colleagues have shown that PQS is able to promote the formation of membrane vesicles off the outer Pseudomonas membrane. Pseudomonas is known to naturally produce these vesicles, however PQS directly induces changes in the lipid membrane to form such vesicles. –
Furthermore, these vesicles have the strong potential to be able to capture a variety of macromolecules that are in the vicinity of the membrane bleb. A prime example would be the beta-lactamases that are in the periplasmic space. But we could imagine other molecules being packaged up and delivered. — Molecules that are packaged could be delivered to the same cells in the population, or to foreign cells of different species, perhaps even to humans during pathogenesis. The possibilities are endless as to what these membrane vesicles could be providing instructions for. — One very important piece of data that is missing from this model, is that researchers have yet to observe membrane vesicles of one cell fusing with another cell. This would provide solid evidence that these vesicles could, in fact, deliver signals to other cells.”
See the 2005 publication Membrane vesicles traffic signals and facilitate group activities in a prokaryote: “Many bacteria use extracellular signals to communicate and coordinate social activities, a process referred to as quorum sensing1. Many quorum signals have significant hydrophobic character, and how these signals are trafficked between bacteria within a population is not understood. Here we show that the opportunistic human pathogen Pseudomonas aeruginosa packages the signalling molecule 2-heptyl-3-hydroxy-4-quinolone
(pseudomonas quinolone signal; PQS)2 into membrane vesicles that serve to traffic this molecule within a population. Removal of these vesicles from the bacterial population halts cell–cell communication and inhibits PQS-controlled group behaviour. We also show that PQS actively mediates its own packaging and the packaging of other antimicrobial quinolines produced by P. aeruginosa into vesicles. These findings illustrate that a prokaryote possesses a signal trafficking system with features common to those used by higher organisms and outlines a novel mechanism for delivery of a signal critical for coordinating group behaviour in P. aeruginosa.”
Clinicaltrials.gov shows four clinical trials related to QS, two completed, one terminated and one recruiting.
A few final words
The worlds of quorum sensing and quorum sensing inhibition appears to be just opening up and indeed may offer a powerful new set of weapons against infectious diseases. I am left with a few questions that may be reflective only of my ignorance or of the early state of research in this area.
To what extent are the QS inhibitors generic across multiple species, and to what extent are they species–specific? A great many of the studies of QS inhibitors have focused on QS inhibition in only one or two species, namely Chromobacterium violaceum and Pseudomonas aeruginosa. This is well and good but leaves open the question of whether the substances equally inhibit QS in a host of other pathogens.
To the extent that the QS inhibitors are generic, what are the implications of their use in vivo, given that they might also inhibit QS in beneficial bacteria?
To what extent is the inhibition of quorum sensing the main mechanism of phytochemicals for combating infectious diseases? What other companion mechanisms may be involved?
What are the extra-cellular vehicles for bacterial QS communications? Do they mainly involve free-floating signal molecules activating cell surface receptors, or are these signaling molecules frequently packaged into vesicles, perhaps even activating bacterial cells via endocytosis? If vesicles are involved, do they commonly carry multiple signaling molecules? I do know that in the case of human QS cell communications, exosome packaging may be involved.
There is much more to be said about quorum sensing and I will continue my discussion of this topic in the Quorum sensing Part 2 blog entry.
Life depends on molecular messaging. Cells and organisms – simple ones like c-elegans and complicated ones like humans – are constantly exchanging internal molecular messages. These messages are critical for articulating development, tissue homeostasis, repairs of various kinds, fighting infectious bacteria and other essential functions of life, Members of different species – including us humans – use molecular messages to sense and gain information about members of other species, to communicate with them, and to jockey for a survival edge with them. We sometimes use such messaging to fool or blindside each other. This applies to bacteria-bacteria interactions, to bacteria-plant interactions, to bacteria-animal interactions and to all combinations of the above. This blog entry covers recent research regarding such forms of messaging
The Quorum Sensing Part 1 blog entry described how many species of bacteria use quorum sensing (QS) molecular messaging to determine the best survival strategy for members of that species under particular conditions, and to drive implementation actions. Moreover, that blog entry discussed how blocking of quorum sensing through use of phytochemicals could be an effective antibacterial strategy that might work well against colony-forming antibiotic resistant bacterial strains. The focus was on intra-species communications. This blog entry, instead, focuses on molecular communications and quorum sensing in inter-species communications and among animal and human cells.
Topics that will be touched on include:
The importance of molecular packing of signaling molecules into vesicles and exosomes,
The roles of QS in biofilm formation,
Molecular messaging of cells in response to infection and stresses,
The role of QS in wound healing, and
The roles of QS in periodontal disease and the formation of tooth cavities
On inter-species molecular communications among biological organisms
Biological organisms may use a wide variety of means for communication including visual, sound, tactical, and olfactory signals, and languages of many kinds. A chameleon can change his color, and octopus can change his spots, bees can engage in flight acrobatics to signal danger, flowers can use bright colors and light-polarizing molecules to attract bees for pollination, humans can shout at each other or write treatises like this one. The list of communications strategies used by biological entities goes on and on.
This blog entry is concerned with molecular communications. Some examples are very familiar, for example a skunk squirting out butyl mercaptan, a very smelly substance, when the skunk senses danger. Members of many species emit and sense pheromones, chemical messengers addressed to members of the same species that can affect social behavior such as the presence of danger or opportunities for sexual engagement. Ants leave trails of chemical messengers to delineate pathways to food. Here, I am concerned with less-familiar molecular communications that originate at the cell level and that may affect health and longevity. Some of the key things I will be discussing are:
Molecular communications in many instances take place through transmission of molecules packaged into microcapsules known as vesicles or exosomes.
Quorum’s sensing enters in for both intra-and inter-species communications when cells act in concert, such as Staphylococcus bacteria in an infection deciding to form a biofilm or release virulence factors, or when immune system cells coordinate a defense.
The single-pathogen disease model is inadequate In many real-world infectious disease situations in humans, such as a case of wound healing, of gum infection of lung infection and chronic rhinosinusitis, Instead, grasping what is going on and identifying an effective therapy requires understanding that pathogens exist in multi-species communities that form biofilms. In such a community, multiple species of bacteria and microflora as well as human cells communicate and exercise quorum sensing among each other and the net result is determined by complex signaling interactions.
The concepts treated here, of exosome communications, of quorum sensing, of rich interspecies mutual involvement and communications, are important for lending new understanding in many areas related to infectious diseases, including wound healing,chronic rhinosinusitis, lung infections, and periodontal disease.
It is very interesting that the publications which discuss exosome communications in disease processes rarely if ever mentioning quorum sensing while the publications which discuss quorum sensing in disease processes rarely mention exosome communications. I assume that this reflects the fact that different communities of researchers are involved in exosome communications and quorum sensing and that these two bodies of researchers probably don’t communicate with each other very well. I try to combine knowledge from these two areas of research together in this blog entry.
Molecular communications from a cell may be via direct release of messenger proteins into the cell’s immediate environment or via nano-sized vesicles, particularly exosomes, which enclose molecular signaling cargo within protective membranes.
The 2011 publication Microvesicles: Intercellular Vectors of Biological Messages relates”Cells communicate directly by cell-cell contact and indirectly via the release of mediators. But gaining a greater appreciation recently is the identification and characterization of intercellular communication through the secretion of microvesicles (MVs). MVs—small vesicles that comprise microparticles (MPs) and exosomes (1) —released from a wide variety of cells, can be considered micro-messengers. Whereas exosomes are released into the extracellular compartment by exocytosis, MPs are shed from the blebbing plasma membrane, and the composition and effects of both on target cells differ depending on the cell from which originate and the type of stimulus involved in their formation. MVs are obtained after several steps of centrifugation, whereby MPs are defined as the MVs obtained by centrifugation at < 100,000 × g, whereas exosomes are isolated by centrifugation at > 100,000 × g. Because the processes by which exosomes and MPs work are different, some researchers have claimed that such broad observations impede the defining and understanding of MV actions (2). — MPs are small vesicles, heterogeneous in size [(0.05–1 μm, a characteristic which is often used to distinguish MPs from exosomes (< 0.1 μm) and platelets (> 1 μm), respectively] and composition, with pro-coagulant and pro-inflammatory properties. Although all cell types can theoretically release MPs, the determination of the their origins has established that MPs can be released from the plasma membrane of circulating cells (such as platelets, erythrocytes, T and B cells, and monocytes); cells from the vascular wall (endothelial and smooth muscle cells); and tumor cells (3, 4). Even if the mechanisms governing MP formation are complex and not well understood, it is well appreciated that cell activation and apoptosis are the two main cellular processes that lead to their formation (Figure 1). — Exosomes are small, natural membrane vesicles released by a wide variety of cell types into the extracellular compartment by exocytosis. Exosomes were described initially during the secretion of MVs of endocytic origin by reticulocytes (8). Indeed, exosomes are formed within endosomes by invagination of the limiting membranes, resulting in the formation of multivesicular bodies. Subsequently, multivesicular bodies fuse with the plasma membrane and release exosomes into the extracellular environment (9). Secretion of exosomes can be spontaneous or induced depending on the cell type. Reticulocytes, T cells, mastocytes, and resting B cells secrete detectable levels of exosomes following the activation of a cell surface receptor. In contrast, dendritic cells, macrophages, and epithelial cells constitutively secrete exosomes in vitro, as do most tumor cells (10). The identification of MVs as exosomes is based on morphological and biochemical criteria. Thus, exosomes are obtained after high speed centrifugation (10); to confirm their presence and purity, immunoblotting analysis with antibody directed against exosomal markers like tetraspanins, heat shock protein (HSP)70, HSP90, or elongation factor-1α are needed. Also, electron microscopy or flow cytometry may be used for detection of exosomes, although there are claims that exosomes would be too small to be detected by flow cytometry. — Although exosomes do not contain any proteins from nuclear, mitochondrial, endoplasmic reticulum or Golgi apparatus, as a consequence of their endosomal origin, they are also selectively enriched in mRNA and miRNA, allowing genetic exchange between cells (11). Because MVs “hijack” the cytoplasm, capturing cytoplasm components and miRNA prior to release from the cell, the packaging of miRNAs in the cell may be random (12). — The mechanisms by which MPs and exosomes transfer biological information to recipient cells are not completely understood, but they may involve direct interaction with surface-expressed ligands or the transfer of surface receptors, proteins, mRNA, miRNA, and bioactive lipids. Moreover, they may serve as vehicles to transfer infectious particles (e.g., “Trojan horse” mechanisms) and to deliver intact organelles. Up to now, four mechanisms (ligand-receptor interaction, transfer, fusion, and internalization) by which information is transmitted by MVs have been reported (Figure 1, Table 1).”
“Figure 1 Schematic representation of mechanisms for the formation of microparticles and exosomes and the different ways by which they interact with target cells Once microparticles (top) and exosomes (bottom) are released, they can interact with target cells through four mechanisms: (1) ligand/receptor interaction, (2) protein transfer, (3) membrane fusion, or (4) internalization.
Exosome packaging of molecular messages offers a number of advantages including protection of signaling cargo from being degraded due to factors in the extra-cellular medium and an ability to target the cargo messages to particular cells.
There is a lot more to be said about exosomeshow and when they are produced and what they do. Besides being conveyors of signaling molecules, exosomes can perform other functions such as ridding cells of lipofuscin trash. My colleague Jim Watson has been studying them recently and is likely to generate one or more blog entries on them in the near future. I mention them here because of their critical role in molecular inter and intra-species communications.
When the Japanese attacked Pearl Harbor, it was a clear message of war One form of bacterial interspecies messaging is an attack. Pathogenic bacteria and parasites can use exosomes to transport virulence factors into cells of a host organism and infect them.
The 2010 e-publication An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages reported: “Specialized secretion systems are used by numerous bacterial pathogens to export virulence factors into host target cells. Leishmania and other eukaryotic intracellular pathogens also deliver effector proteins into host cells; however, the mechanisms involved have remained elusive. In this report, we identify exosome-based secretion as a general mechanism for protein secretion by Leishmania, and show that exosomes are involved in the delivery of proteins into host target cells. Comparative quantitative proteomics unambiguously identified 329 proteins in Leishmania exosomes, accounting for >52% of global protein secretion from these organisms. Our findings demonstrate that infection-like stressors (37 degrees C +/- pH 5.5) upregulated exosome release more than twofold and also modified exosome protein composition. Leishmania exosomes and exosomal proteins were detected in the cytosolic compartment of infected macrophages and incubation of macrophages with exosomes selectively induced secretion of IL-8, but not TNF-alpha. We thus provide evidence for an apparently broad-based mechanism of protein export by Leishmania. Moreover, we describe a mechanism for the direct delivery of Leishmania molecules into macrophages. These findings suggest that, like mammalian exosomes, Leishmania exosomes function in long-range communication and immune modulation.”
Exosome release of virulence factors appears to be a general property of mycobacterial species
The 2011 publication Mycobacteria release active membrane vesicles that modulate immune responses in a TLR2-dependent manner in mice speaks to a similar point: “Bacteria naturally release membrane vesicles (MVs) under a variety of growth environments. Their production is associated with virulence due to their capacity to concentrate toxins and immunomodulatory molecules. In this report, we show that the 2 medically important species of mycobacteria, Mycobacterium tuberculosis and Mycobacterium bovis bacille Calmette-Guérin, release MVs when growing in both liquid culture and within murine phagocytic cells in vitro and in vivo. We documented MV production in a variety of virulent and nonvirulent mycobacterial species, indicating that release of MVs is a property conserved among mycobacterial species. Extensive proteomic analysis revealed that only MVs from the virulent strains contained TLR2 lipoprotein agonists. The interaction of MVs with macrophages isolated from mice stimulated the release of cytokines and chemokines in a TLR2-dependent fashion, and infusion of MVs into mouse lungs elicited a florid inflammatory response in WT but not TLR2-deficient mice. When MVs were administered to mice before M. tuberculosis pulmonary infection, an accelerated local inflammatory response with increased bacterial replication was seen in the lungs and spleens. Our results provide strong evidence that actively released mycobacterial vesicles are a delivery mechanism for immunologically active molecules that contribute to mycobacterial virulence. These findings may open up new horizons for understanding the pathogenesis of tuberculosis and developing vaccines.” Other publications listed below describe how infected macrophages are the ones which alert the immune system and promote the inflammatory response. They also do this via a system of exosome communications, in this case intra-species.”
Animal and human cells stressed by bacterial or parasitic infection or other stressors generate exosome communications to alert other cells of their plight and to initiate responsive actions. Stressed cells at an injury site generate signals to immune system cells calling for an inflammatory response. In particular, macrophages may emit exosomes containing chemokines, cytokines and chemotaxic factors that initiate neutrophil, macrophage, and dendritic cell migration to the injury site.
A number of recent publications have been addressed to this point. I listed a few examples. Later, I will discuss how signaling from infectious bacteria in biofilm colonies can interfere with or inhibit healing human exosome communications.
The 2007 publication Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo relates “Intracellular pathogens and the molecules they express have limited contact with the immune system. Here, we show that macrophages infected with intracellular pathogens Mycobacterium tuberculosis, M bovis BCG, Salmonella typhimurium, or Toxoplasma gondii release from cells small vesicles known as exosomes which contain pathogen-associated molecular patterns (PAMPs). These exosomes, when exposed to uninfected macrophages, stimulate a proinflammatory response in a Toll-like receptor– and myeloid differentiation factor 88–dependent manner. Further, exosomes isolated from the bronchoalveolar lavage fluid (BALF) of M bovis BCG–infected mice contain the mycobacteria components lipoarabinomannan and the 19-kDa lipoprotein and can stimulate TNF-α production in naive macrophages. Moreover, exosomes isolated from M bovis BCG– and M tuberculosis–infected macrophages, when injected intranasally into mice, stimulate TNF-α and IL-12 production as well as neutrophil and macrophage recruitment in the lung. These studies identify a previously unknown function for exosomes in promoting intercellular communication during an immune response to intracellular pathogens, and we hypothesize that extracellular release of exosomes containing PAMPs is an important mechanism of immune surveillance.”
A 2013 publication Microparticles from mycobacteria-infected macrophages promote inflammation and cellular migration reports: “Mycobacterium tuberculosis infection is characterized by a strong inflammatory response whereby a few infected macrophages within the granuloma induce sustained cellular accumulation. The mechanisms coordinating this response are poorly characterized. We hypothesized that microparticles (MPs), which are submicron, plasma membrane-derived vesicles released by cells under both physiological and pathological conditions, are involved in this process. Aerosol infection of mice with M. tuberculosis increased CD45(+) MPs in the blood after 4 wk of infection, and in vitro infection of human and murine macrophages with mycobacteria enhanced MP release. MPs derived from mycobacteria-infected macrophages were proinflammatory, and when injected into uninfected mice they induced significant neutrophil, macrophage, and dendritic cell recruitment to the injection site. When incubated with naive macrophages, these MPs enhanced proinflammatory cytokine and chemokine release, and they aided in the disruption of the integrity of a respiratory epithelial cell monolayer, providing a mechanism for the egress of cells to the site of M. tuberculosis infection in the lung. In addition, MPs colocalized with the endocytic recycling marker Rab11a within macrophages, and this association increased when the MPs were isolated from mycobacteria-infected cells. M. tuberculosis-derived MPs also carried mycobacterial Ag and were able to activate M. tuberculosis-specific CD4(+) T cells in vivo and in vitro in a dendritic cell-dependent manner. Collectively, these data identify an unrecognized role for MPs in host response against M. tuberculosis by promoting inflammation, intercellular communication, and cell migration.”
A May 2013 article Immunomodulatory impact of leishmania-induced macrophage exosomes: a comparative proteomic and functional analysis reports: “Released by many eukaryotic cells, the exosomes are 40–100 nm vesicles shown to operate over the complex processes of cell-cell communication. Among the metazoan cell lineages known to generate exosomes is the mononuclear phagocyte lineage, a lineage that parasites such as Leishmania are known to subvert as host cells. We previously reported that mouse macrophage signaling and functions are modified once co-incubated with exoproteome of Leishmania promastigotes. Using mass spectrometry analysis, we were curious to further compare the content of purified exosomes released by the J774 mouse macrophage cell line exposed or not to either LPS or to stationary phase Leishmania mexicana promastigotes. Collectively, our analyses resulted in detection of 248 proteins, ~50–80% of which were shared among the three sources studied. Using exponentially modified protein abundance index (emPAI) and network analyses, we found that the macrophage exosomes display unique signatures with respect to composition and abundance of many functional groups of proteins, such as plasma membrane-associated proteins, chaperones and metabolic enzymes. Moreover, for the first time, L. mexicana surface protease GP63 is shown to be present in exosomes released from J774 macrophages exposed to stationary phase promastigotes. We observed that macrophage exosomes are able to induce signaling molecules and transcription factors in naive macrophages. Finally, using qRT-PCR, we monitored modulation of expression of multiple immune-related genes within macrophages exposed to exosomes. We found all three groups of exosomes to induce expression of immune-related genes, the ones collected from macrophages exposed to L. mexicana sharing properties with exosomes collected from macrophage left unexposed to any agonist. Overall, our results allowed depicting that protein sorting into macrophage-derived exosomes depends upon the cell status and how such distinct protein sorting can in turn impact the functions of naive J774 cells. — Author Summary: Secreted vesicles, such as exosomes, are now considered as an important route of communication among eukaryotic cells. Depending on the donor cell source and protein content, these vesicles are expected to distinctly impact the recipient cell properties. Here, three groups of exosomes released by the mouse macrophage cell line J774 exposed or not – naive exosomes – to either Leishmania mexicana promastigotes or to LPS were compared through proteomic analysis. Also, their biological activities on naive J774 macrophages were tested. Regardless of the source, the three groups of exosomes shared 50–80% of their proteins, although their relative abundances differed, especially those associated with the plasma membrane. Post exposure to one out of the three groups of exosomes, naive J774 recipient macrophages were compared for their profile of immune transcripts. Of note, whether they were exposed to either naive exosomes or to L. mexicana- induced exosomes, the naive J774 macrophages shared similar immune transcriptional signatures, the latter being distinct from the ones displayed by the macrophages exposed to LPS-induced exosomes. These data are discussed within the context of the unique cross talk that accounts for the early establishment of an immunomodulatory parasite such as Leishmania in its mammalian host.”
Not only can macrophages generate signaling molecules when stressed, but also signaling molecules in exosomes can directly influence the behavior of macrophages.
The 2011 publication [A preliminary study on the influence of human plasma exosomes-like vesicles on macrophage Wnt5A-Ca²+ pathway] reports: “OBJECTIVE: To study the influence of human plasma exosomes-like vesicles on the regulatory function of macrophages. METHODS: The exosomes-like vesicles were purified from healthy donors plasma with a series of high-speed centrifugation and ultrafiltration. Macrophages were derived from cultured human blood monocytes. The molecular markers of macrophages were assayed by FACS. After cultured with exosomes-like vesicles, the changes of macrophages cytoplasma Ca(2+), and related genes and proteins were assayed by FACS, RT-PCR and Western Blot, respectively. RESULTS: After cultured with exosomes-like vesicles, mean fluorescent intensity (MFI) of macrophages cytoplasma Ca(2+) was increased. The vesicles enhanced macrophages to express cytokines genes, the expression of IL-1β and TNF-α genes being increased by 0.85 and 1.69 times respectively at 2 h, and that of IL-6 gene 3.7 times compared with the control at 8 h. However, the vesicles inhibited the expression of macrophages IL-10 gene, had no influence on the Frizzled5 receptor expression and could induce CaMKII phosphorylation. CONCLUSIONS: Exosomes-like vesicles can up-regulate macrophages expression of inflammatory cytokines genes, and increase the secretion of inflammatory cytokines by activating the Wnt5A-Ca(2+) signaling pathway.”
Exosomes from infected macrophages may contain many different proteins including antigenic (antibody-producing) ones. The 2010 publication Proteomic analysis identifies highly antigenic proteins in exosomes from M. tuberculosis-infected and culture filtrate protein-treated macrophages relates: “Exosomes are small 30-100 nm membrane vesicles released from hematopoietic and nonhematopoietic cells and function to promote intercellular communication. They are generated through fusion of multivesicular bodies with the plasma membrane and release of interluminal vesicles. Previous studies from our laboratory demonstrated that macrophages infected with Mycobacterium release exosomes that promote activation of both innate and acquired immune responses; however, the components present in exosomes inducing these host responses were not defined. This study used LC-MS/MS to identify 41 mycobacterial proteins present in exosomes released from M. tuberculosis-infected J774 cells. Many of these proteins have been characterized as highly immunogenic. Further, since most of the mycobacterial proteins identified are actively secreted, we hypothesized that macrophages treated with M. tuberculosis culture filtrate proteins (CFPs) would release exosomes containing mycobacterial proteins. We found 29 M. tuberculosis proteins in exosomes released from CFP-treated J774 cells, the majority of which were also present in exosomes isolated from M. tuberculosis-infected cells. The exosomes from CFP-treated J774 cells could promote macrophage and dendritic cell activation as well as activation of naïve T cells in vivo. These results suggest that exosomes containing M. tuberculosis antigens may be alternative approach to developing a tuberculosis vaccine.”
A pathogen can also sometimes trick host cells into sending microvesicle or exosome messages that actually help the pathogen.
The 2013 publication Interplay of host-pathogen microvesicles and their role in infectious disease relates “The release of extracellular vesicles, whether MVs (microvesicles) or exosomes, from host cells or intracellular pathogens is likely to play a significant role in the infection process. Host MVs may fuse with pathogen surfaces to deliver host complement regulatory proteins. They may also deliver cytokines that enhance invasion. Decoy functions are also possible. Whereas host MVs may direct pathogens away from their target cells, pathogen MVs may in turn redirect complement membrane-attack complexes away from their target pathogen. An understanding of the mechanisms of this interplay, bringing about both immune evasion and enhanced invasion, will help to direct future research with a view to rendering pathogens more susceptible to immune attack or in improving drug efficacy. It should also be possible to use MVs or exosomes isolated directly from the pathogens, or from the cells infected with pathogens, to provide alternative vaccination strategies.”
The 2011 publication Exosomes released from M. tuberculosis infected cells can suppress IFN-γ mediated activation of naïve macrophages describes another situation where human macrophage cell exosome communications are negatively affected by a pathogen so as to compromise immune system response. “BACKGROUND: Macrophages infected with Mycobacterium tuberculosis (M.tb) are known to be refractory to IFN-γ stimulation. Previous studies have shown that M.tb express components such as the 19-kDa lipoprotein and peptidoglycan that can bind to macrophage receptors including the Toll-like receptor 2 resulting in the loss in IFN-γ responsiveness. However, it is unclear whether this effect is limited to infected macrophages. We have previously shown that M.tb-infected macrophages release exosomes which are 30-100 nm membrane bound vesicles of endosomal origin that function in intercellular communication. These exosomes contain mycobacterial components including the 19-kDa lipoprotein and therefore we hypothesized that macrophages exposed to exosomes may show limited response to IFN-γ stimulation. METHODOLOGY/PRINCIPAL FINDINGS: Exosomes were isolated from resting as well as M.tb-infected RAW264.7 macrophages. Mouse bone marrow-derived macrophages (BMMØ) were treated with exosomes +/- IFN-γ. Cells were harvested and analyzed for suppression of IFN-γ responsive genes by flow cytometry and real time PCR. We found that exosomes derived from M.tb H37Rv-infected but not from uninfected macrophages inhibited IFN-γ induced MHC class II and CD64 expression on BMMØ. This inhibition was only partially dependent on the presence of lipoproteins but completely dependent on TLR2 and MyD88. The exosomes isolated from infected cells did not inhibit STAT1 Tyrosine phosphorylation but down-regulated IFN-γ induced expression of the class II major histocompatibility complex transactivator; a key regulator of class II MHC expression. Microarray studies showed that subsets of genes induced by IFN-γ were inhibited by exosomes from H37Rv-infected cells including genes involved in antigen presentation. Moreover, this set of genes partially overlapped with the IFN-γ-induced genes inhibited by H37Rv infection. CONCLUSIONS: Our study suggests that exosomes, as carriers of M.tb pathogen associated molecular patterns (PAMPs), may provide a mechanism by which M.tb may exert its suppression of a host immune response beyond the infected cell.”
Other publications that treat host-cell exosome messaging in response to infection or other stressors include:
Again, I saw no mention of quorum sensing while looking at the above exosome-related publications. Later I will cite publications showing that quorum sensing is deeply involved.
Immune system cells likely use quorum sensing for a number of purposes, for example establishing homeostasis of numbers of T cells in a niche.
The 2012 e-publication Quorum-Sensing in CD4(+) T Cell Homeostasis: A Hypothesis and a Model reports “Homeostasis of lymphocyte numbers is believed to be due to competition between cellular populations for a common niche of restricted size, defined by the combination of interactions and trophic factors required for cell survival. Here we propose a new mechanism: homeostasis of lymphocyte numbers could also be achieved by the ability of lymphocytes to perceive the density of their own populations. Such a mechanism would be reminiscent of the primordial quorum-sensing systems used by bacteria, in which some bacteria sense the accumulation of bacterial metabolites secreted by other elements of the population, allowing them to “count” the number of cells present and adapt their growth accordingly. We propose that homeostasis of CD4(+) T cell numbers may occur via a quorum-sensing-like mechanism, where IL-2 is produced by activated CD4(+) T cells and sensed by a population of CD4(+) Treg cells that expresses the high-affinity IL-2Rα-chain and can regulate the number of activated IL-2-producing CD4(+) T cells and the total CD4(+) T cell population. In other words, CD4(+) T cell populations can restrain their growth by monitoring the number of activated cells, thus preventing uncontrolled lymphocyte proliferation during immune responses. We hypothesize that malfunction of this quorum-sensing mechanism may lead to uncontrolled T cell activation and autoimmunity. Finally, we present a mathematical model that describes the key role of IL-2 and quorum-sensing mechanisms in CD4(+) T cell homeostasis during an immune response.”
“Quorum-sensing. The presence of IL-2 and the ability of the Treg cells to detect its levels are crucial to the homeostasis of the immune system. Quorum-sensing in this case is defined as an indirect feedback loop where the IL-2 produced by a subpopulation of activated T cells (among others) is detected (sensed) by a subpopulation of CD4+ Treg cells expressing the high-affinity IL-2Rα-chain; these cells contribute to controlling the number of CD4+ T cells. In other words, the overall CD4+ T cell populations sense the produced quantities of IL-2 and adapt their behavior accordingly. (B) Failure of quorum-sensing by defective sensor molecule. The inability to detect IL-2 because of defects in IL-2R expression (in IL-2Rα−/− or IL-2Rβ−/− mice) or signaling (in STAT5−/− mice) leads to lymphoid hyperplasia and autoimmune disease. (C) Failure of quorum-sensing due to absence of the sensed molecule. In the absence of IL-2, Treg cells do not survive, which causes lymphoid hyperplasia and autoimmune pathology.”
Many forms of human bacterial pathologies – whether they be in the lung, in the mouth, or in nonhealing wounds – involve biofilms in which multiple species of bacteria cooperate for their mutual benefit and there is a rich network of quorum sensing communications.
The 2008 publication Quorum Sensing and Bacterial Social Interactions in Biofilms relates: “ — The ability of bacteria to communicate and behave as a group for social interactions like a multi-cellular organism has provided significant benefits to bacteria in host colonization, formation of biofilms, defense against competitors, and adaptation to changing environments. Importantly, many QS-controlled activities have been involved in the virulence and pathogenic potential of bacteria. Therefore, understanding the molecular details of quorum sensing mechanisms and their controlled social activities may open a new avenue for controlling bacterial infections.”
It is time to get beyond looking at infections in terms of single pathogens.
Continuing (ref) “ — It was believed for many years that bacteria, unlike eukaryotic organisms, behaved as self-sufficient individuals and maintained a strictly unicellular life-style [1–3]. During infections, bacterial mass was considered nothing more than the sum of these individuals. Our perception of bacteria as unicellular life-style was deeply rooted in the pure culture paradigm of Robert Koch’s era, when Koch established his “golden criteria” to define a bacterial pathogen by using pure-culture approaches [3]. Indeed, Koch’s concept has led to the great success in the identification of bacterial pathogens and development of antibiotic treatments in acute bacterial infections [3,4]. However, pure-culture planktonic growth of bacteria rarely exists in natural environments. In fact, bacteria in Nature largely reside in a complex and dynamic surface-associated community called a biofilm [3,5,6]. If viewing an intact biofilm under microscope, one may immediately find that bacteria in biofilms do not randomly stick together, but rather form a well-organized community with numerous specialized configurations [5,6]. One may also find that bacterial cells in biofilms physically interact with each other and maintain ‘intimate’ relationships [5,6]. Even without physical contact, bacteria living at the same community likely secrete small extra-cellular molecules to interact with each other [7–10]. It was not until the last three decades that our view of self-sufficient unicellular lifestyle of bacteria has changed. The advances from at least two major research areas, biofilm development and bacterial quorum sensing, have led us to begin to appreciate, in much more detail for the first time, the concept that bacteria can organize into groups, form well-organized communities, and communicate for coordinated activities or social life that was once believed to be restricted to multi-cellular organisms [3,6–13]. — Microbiologists have discovered an unexpectedly high degree of coordinated multi-cellular behaviors that have led to the perception of biofilms as “cities” of microorganisms [6]. Especially, many bacteria have been found to regulate diverse physiological processes and group activities through a mechanism called quorum sensing, in which bacterial cells produce, detect and respond to small diffusible signal molecule [7–14]. It has long been known that in infectious diseases the invading bacteria need to reach a critical cell density before they express virulence and overwhelm the host defense mechanisms before they initiate an infectious disease [1–3]. Since quorum-sensing mechanisms are widespread in both prokaryotic and single-celled eukaryotic organisms such as fungi [7–9,15], it is not surprising that cell-cell communication through quorum sensing has important implications in microbial infections. A growing body of excellent reviews has highlighted.”
Biofilms are involved in many important infectious disease processes, if not most of them. Studied examples include infectious kidney stones, bacterial endocarditis, cystic fibrosis lung infections, non-healing wounds, chronic rhinosinusitis, periodontal disease and dental caries.
“Four possible mechanisms of biofilm antibiotic resistance. The image is a cross section of a biofilm with the attachment surface (gray) at the bottom and the aqueous phase containing the antibiotic (yellow) at the top. In zones where there is nutrient depletion (red), antibiotic action may be antagonized. Some bacteria may activate stress responses (green), while others may differentiate into a protected phenotype (purple).”
Biofilms occur naturally in nature, such as in the beds of streams, and some are beneficial to humans. You can find out all about biofilms in a nice tutorial here. You can also check out an earlier blog entry Chronic rhinosinusitis, MRSA, biofilms and manuka honey.
Because of molecular communications, there are no true bystander cells when it comes to infectious attacks.
The “bystander effect” has long puzzled biologists. You do something to some cells like expose them to radiation, and the impact definitely shows up in not only the irradiated cells but also in neighboring cells that did not get irradiated. But the fact is that because of molecular communications from the irradiated cells, the others are affected too. This bystander effect effect shows up in many domains of biology, including attacks by infectious bacteria.. For example, the 2011 publication Mannosylated lipoarabinomannan balances apoptosis and inflammatory state in mycobacteria-infected and uninfected bystander macrophages. “To assess the role of mannosylated lipoarabinomannan (ManLAM) in the inflammatory and apoptotic response of mycobacteria-infected and uninfected, bystander cells we applied a mouse macrophage model of infection with avirulent strains–Mycobacterium bovis BCG, Mycobacterium tuberculosis (MTB) H37Ra and compared with a virulent MTB H37Rv strain infection. ManLAM contributed to the infection of macrophages by protection from apoptosis with stabilized Bcl-2 expression and down-regulated Bax expression for infected cells (BCG) or with stabilized Bcl-2 expression for uninfected bystander target cells (H37Ra). Additionally, ManLAM up-regulated FasL expression on the infected cells. Active extracellular signal-regulated kinase (ERK1/2) in BCG and H37Rv infection provided an anti-apoptotic effect by stabilization of anti-apoptotic Bcl-2 expression in the infected cells. Inhibitors specific for c-Jun-NH2-terminal kinase or stress-activated kinase (JNK) and p38 kinase decreased apoptosis of infected cells (BCG, H37Ra) and of uninfected bystanders (H37Ra) by down-regulating Bax. ManLAM significantly down-regulated production of pro-inflammatory IL-12 and TNF-alpha and activation of JNK by both avirulent strains. We conclude that by stabilization of Bcl-2 expression, down-regulation of JNK activity and down-regulation of pro-inflammatory cytokines production ManLAM can contribute to suppression of apoptosis and inflammatory reaction of uninfected, bystander cells.” In rich networks of biological communications, there are no real bystanders.
I hypothesize that most basic decisions of cells that drive cell behavior and responses are not decisions made by individual cells but are common decisions made by the community of cells involved, triggered by quorum sensing and likely communicated by exosome signaling. These cell communities can include pathogenic bacteria of a given species attacking a wound or organ, human cells of given types such as T cells and stem cell types, microbiota, and plant cells of a given type.
This is a rather sweeping conjecture of my own and I will argue for it by citing examples.
In the Quorum Sensing Part 1 blog entry I have already cited for multiple infectious bacteria how quorum sensing, counting the relative number of bacteria present, drives behavior which determines whether the bacteria lie low, whether they form protective biofilms, or whether they go on the attack and express virulence. These are community decisions and the bacteria tend to act in concert. Publications like ones cited here above suggest that the quorum sensing communications at least partially involve messages encapsulated in exosomes or other microvesicles.
In healthy individuals, T cells make community decisions as to which cells represent “self,” and which ones represent “foreign invader” and should therefore be attacked. The 2012 publication Quorum sensing allows T cells todiscriminate betweenself and nonself reports: “T cells orchestrate pathogen-specific adaptive immune responses by identifying peptides derived from pathogenic proteins that are displayed on the surface of infected cells. Host cells also display peptide fragments from the host’s own proteins. Incorrectly identifying peptides derived from the body’s own proteome as pathogenic can result in autoimmune disease. To minimize autoreactivity, immature T cells that respond to self-peptides are deleted in the thymus by a process called negative selection. However, negative selection is imperfect, and autoreactive T cells exist in healthy individuals. To understand how autoimmunity is yet avoided, without loss of responsiveness to pathogens, we have developed a model of T-cell training and response. Our model shows that T cells reliably respond to infection and avoid autoimmunity because collective decisions made by the T-cell population, rather than the responses of individual T cells, determine biological outcomes. The theory is qualitatively consistent with experimental data and yields a criterion for thymic selection to be adequate for suppressing autoimmunity.”
3. “Bystander” cellular responses are only explainable in terms of this hypothesis. Consider the radiation bystander effect for example. When a hormetic dose of radiation is applied ti human cells, not only do the cells that receive that radiation experience upgrading of their stress responses, but also neighboring cells that themselves received no radiation also get their stress responses upgraded. If the radiation dose is super-hormetic and the irradiated cells experience damage, so will the un-irradiated bystander cells be damaged. This bystander effect might be immune system mediated and it might induce a hormetic response (ref). Also the effect could be a direct result of cell to cell signaling (ref)(ref). In any case, quorum-sensing decision is made by the community of cells and this decision is communicated to all cells, radiated and not-radiated alike. The bystander cells go along with what the community decides. Based on other publications, I surmise that it is highly probable that communication originates with exosomes released from the radiation-stressed cells. Quorum sensing appears to be a necessary mechanism for decisions to be made among all the cells in the community A quorum sensing decision could be 1. to ignore the stress signals, 2. To upgrade their stress responses, 4. To become damaged, or 4. To scream like crazy for the immune system to weigh in and inflammation to be triggered. It appears that cells are very socialist and lemming-like in their behavior. They appear to respond collectively depending on how cells in the community feel at the moment. This is probably one of the reasons why stress responses are dependent on circadian rhythms and not just a function of the stress itself.
Human diseases may entail interspecies molecular communications not only between infectious bacteria of a given species and cells of the host human, but also signaling actions of third-party species organisms including microflora. The conventional picture of a bacterial infection being a contest between attacking bacterial cells and defending human cells can be far too simple. Each of multiple co-located species may use quorum sensing for regulation of gene expression and the net pathological impact may depend critically on messages from seemingly innocuous third-party organisms.
This was pointed out back in 2003 in the publication Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. “The change in gene expression patterns in response to host environments is a prerequisite for bacterial infection. Bacterial diseases often occur as an outcome of the complex interactions between pathogens and the host. The indigenous, usually non-pathogenic microflora is a ubiquitous constituent of the host. In order to understand the interactions between pathogens and the resident microflora and how they affect the gene expression patterns of the pathogens and contribute to bacterial diseases, the interactions between pathogenic Pseudomonas aeruginosa and avirulent oropharyngeal flora (OF) strains isolated from sputum samples of cystic fibrosis (CF) patients were investigated. Animal experiments using a rat lung infection model indicate that the presence of OF bacteria enhanced lung damage caused by P. aeruginosa . Genome-wide transcriptional analysis with a lux reporter-based promoter library demonstrated that 4% of genes in the genome responded to the presence of OF strains using an in vitro system. Characterization of a subset of the regulated genes indicates that they fall into seven functional classes, and large portions of the upregulated genes are genes important for P. aeruginosa pathogenesis. Autoinducer-2 (AI-2)-mediated quorum sensing, a proposed interspecies signalling system, accounted for some, but not all, of the gene regulation. A substantial amount of AI-2 was detected directly in sputum samples from CF patients and in cultures of most nonpseudomonad bacteria isolated from the sputa. Transcriptional profiling of a set of defined P. aeruginosa virulence factor promoters revealed that OF and exogenous AI-2 could upregulate overlapping subsets of these genes. These results suggest important contributions of the host microflora to P. aeruginosa infection by modulating gene expression via interspecies communications.” – “Broad response in P. aeruginosa seems to be induced by the presence of OF. Although competition for nutrients was expected to be one of the factors, the observed response appears to be a result of more complex interactions between P. aeruginosa and OF bacteria. — The upregulation of a significant number of virulence genes in P. aeruginosa by OF strains is intriguing and probably explains the increased lung damage observed in the presence of the OF strain in the animal experiments. The upregulation of a pathogen’s virulence factors by microflora underscores the importance of bacterial interactions in pathogenicity. The interactions between bacteria and their host are believed to determine the evolution of many of the virulence factors that pathogens possess — Bacterial pathogens, especially those that inhabit both environmental niches as well as animal hosts, may rely on these interactions to maintain their virulence when out of the hosts. On the other hand, these virulence factors may play a role in the development of microbial communities. – The regulation of P. aeruginosa gene expression and virulence involves its own repertoire of cell–cell signaling molecules (Fuqua et al., 2001; Miller and Bassler, 2001; Whitehead et al., 2001; Smith and Iglewski, 2003). The pathways through which the OF strains affect P. aeruginosa gene expression are expected to be multifactorial and complex. One of the signals seems to be autoinducer-2 (AI-2) produced by non-pseudomonad strains. It seems that P. aeruginosa is modulating its behaviour by monitoring the environmental conditions and by eavesdropping on the other bacteria via AI-2 and probably other signals. AI-2 in the sputum samples could reach a substantial amount, which is readily detectable by the V. harveyi assay. The regulation of a number of virulence factor genes by AI-2 indicates that this signal produced by OF strains contributed, at least in part, to the observed modulation of P. aeruginosa gene expression and changes in its pathogenicity. The AI-2-regulated P. aeruginosa virulence factor genes partially overlap those modulated by OF strains, suggesting that, in the co-culture experiments, AI-2 was one of the signals but not the only signal produced by the OF that regulates P. aeruginosa gene expression. In the lungs of CF patients, this AI-2-mediated effect could also originate from secondary pathogens, suggesting that the virulence of secondary pathogens could also be delivered through enhancing the virulence of the primary pathogen, P. aeruginosa. It is also possible that P. aeruginosa could influence virulence factor genes of secondary pathogens or those potentially present in the OF strains.”
The 2006 review publication Communication in bacteria: an ecological and evolutionary perspectivecomments on this picture: “Individual bacteria can alter their behaviour through chemical interactions between organisms in microbial communities. This is generally referred to as quorum sensing. Frequently, these interactions are interpreted in terms of communication to mediate coordinated, multicellular behaviour. We show that the nature of interactions through quorum-sensing chemicals does not simply involve cooperative signals, but entails other interactions such as cues and chemical manipulations. These signals might have a role in conflicts within and between species. The nature of the chemical interaction is important to take into account when studying why and how bacteria react to the chemical substances that are produced by other bacteria.”
The 2008 treatise Bio-Communication of Bacteria and their Evolutionary Roots in Natural Genome Editing Competences of Viruses offers a nice overview of bacterial communications systems and their evolutionary origins: “Communicative competences and the use of a semiochemical vocabulary enable bacteria to develop, organize and coordinate rich social life with a great variety of behavioral patterns even in which they organise themselves like multicellular organisms. They have existed for almost four billion years and still survive, being part of the most dramatic changes in evolutionary history such as DNA invention, cellular life, invention of nearly all protein types, partial constitution of eukaryotic cells, vertical colonisation of all eukaryotes, high adaptability through horizontal gene transfer and cooperative
multispecies colonisation of all ecological niches. Recent research demonstrates that these bacterial competences derive from the aptitude of viruses for natural genome editing.
In contrast to a book which would be the appropriate space to outline in depth all communicative pathways inherent in bacterial life in this current article I want to give an overview for a broader readership over the great variety of bacterial bio-communication: In a first step I describe how they interpret and coordinate, what semiochemical vocabulary they share and which goals they try to reach. In a second stage I describe transorganismic communication, i.e. the main categories of sign-mediated interactions between bacterial and non-bacterial organisms, and interorganismic communication, i.e. between bacteria of the same or related species. In a third stage I will focus on intraorganismic communication, i.e. the relationship between bacteria and their obligate settlers, i.e. viruses. We will see that bacteria are important hosts for multiviral colonisation and the virally-determined order of nucleic acid sequences, which has implications for our understanding of the evolutionary history of pre-cellular and cellular life.”
Different species of bacteria in a colony may engage in cross talk communications so as to impact the behavior and virulance of each other.
From the 2005 publication Interactions between Oral Bacteria:Inhibition of Streptococcus mutans Bacteriocin Production by Streptococcus gordonii:“Recent studies have shown that quorum sensing modulates both intra- and interspecies cell-cell communications. For example, there is cross-inhibition by quorum-sensing pheromones between Staphylococcus aureus and Staphylococcus epidermidis (21). S. epidermidis pheromones inhibited the S. aureus agr response, which may explain the predominance of S. epidermidis on the skin. Interaction between Bacillus subtilis and Erwinia carotovora provides another example (5), since the former produces an enzyme (AiiA) that inactivates the E. carotovora autoinducers. Inactivation of the autoinducers by AiiA attenuated the virulence of E. carotovora.”
The two central “usual suspect” bacteria for inhibited wound healing are Staphylococcus aureus (SA) and Pseudomonas aeruginosa (PA)
The 2009 publication Staphylococcalbiofilmsimpairwoundhealingbydelayingreepithelializationinamurinecutaneouswoundmodel reported “Bacterial biofilms have gained increasing visibility in recent years as a ubiquitous form of survival for microorganisms in myriad environments. A number of in vivo models exist for the study of biofilms in the setting of medically relevant implanted foreign bodies. Growing evidence has demonstrated the presence of bacterial biofilms in the setting of a number of chronic wound states including pressure sores, diabetic foot ulcers, and venous stasis ulcers. Here we present a novel murine cutaneous wound system that directly demonstrates delayed reepithelialization caused by the presence of a bacterial biofilm. We established biofilms using either Staphylococcus aureus or Staphylococcus epidermidis in splinted cutaneous punch wounds created on the backs of normal C57Bl6/J mice. Wound reepithelialization was significantly delayed by bacterial biofilms. This effect was specifically dependent on the ability of the bacteria to form biofilms as demonstrated by exogenous administration of biofilm inhibiting peptides and the use of mutant Staphylococcus spp. deficient in biofilm formation. This represents the first direct evidence for the effect of bacterial biofilms on cutaneous wound healing. — Biofilms are complex communities of microorganisms associated with surfaces and are characterized by high cell densities associated with an extracellular polymer matrix. In recent years, microbial research has revealed that the bacterial biofilms are far more ubiquitous than previously realized. Indeed, when compared with the traditional view of bacteria growing in planktonic culture with essentially unlimited energy sources, it has become clear that biofilms represent a far more realistic representation of bacterial behavior outside of the laboratory setting.1–5 The genetic signals involved in biofilm formation have been elucidated in several bacterial species of clinical interest, including the Gram-positive cocci Staphylococcus aureus and Staphylococcus epidermidis and the concept of quorum sensing has emerged as an essential means of bacterial communication essential to form the biofilm community.6–12 It is now appreciated that there exist discrete signaling molecules that both promote biofilm assembly as well as biofilm dissolution to facilitate planktonic release of bacteria into another niche.”
The 2012 publication Impact of Pseudomonas aeruginosa quorum sensing on cellular wound healing responses in vitro relates “The virulent capacity of Pseudomonas aeruginosa can largely be ascribed to quorum sensing, i.e. the ability to evade host defence by a coordinated production and secretion of virulence factors. When P. aeruginosa is harboured in chronic wounds, a non-healing condition is often observed. In this study, we examined the in vitro cellular responses of the major cell types of re-epithelialization to supernatants of P. aeruginosa wild-type or an isogenic mutant not expressing quorum sensing-regulated virulence genes. We observed impairment of cell migration in keratinocytes (p = 0.009) and fibroblasts (p = 0.043) when supplementing medium with 20% P. aeruginosa culture supernatants. Cell proliferation was not significantly reduced, except for keratinocytes (p = 0.040). Data show compliance with in vivo observations of proliferating, non-motile epithelial cell behaviour in bacterially contaminated chronic wounds. Our findings suggest that quorum sensing may serve as an interesting target for controlling P. aeruginosa virulence in modern wound care.”
Quorum sensing communications from PA also affect expression in human cells.
The publication The Pseudomonas aeruginosa N-Acylhomoserine Lactone Quorum Sensing Molecules Target IQGAP1 and Modulate Epithelial Cell Migrationrelates “The human pathogen Pseudomonas aeruginosa and other bacteria communicate with each other using quorum sensing (QS). This is important for their growth, virulence, motility and the formation of biofilms. Furthermore, eukaryotic cells “listen and respond” to QS signaling, but the exact mechanisms and receptors on mammalian cells have not been identified.– We suggest that recognition of IQGAP1 by 3O-C12-HSL is a very early event in the communication between bacteria and human epithelial cells.”
Wound healing is a complex process requiring much signaling among human cells. Impaired wound healing can result from colonization of the wound by multiple biofilm-forming bacteria, not just Staphylococcus aureus and Pseudomonas aeruginosa, and the results of their communications.The process can be far from simple, involving simultaneous quorum sensing cross-talk among several species.
The treatise Understanding the effects of bacterial communities and biofilms on wound healing characterizes the complexity involved. “For survival and reproductive success, species of bacteria often rely on close relationships with other species. A collection of bacteria occupying the same physical habitat is called a ‘community’, and one example of a community of micro-organisms is a ‘biofilm’. Biofilms have been implicated in numerous chronic infections including cystic fibrosis, otitis media and prostatitis. Through interactions within a biofilm, the resident population of bacteria is likely to benefit from increased metabolic efficiency, substrate accessibility, enhanced resistance to environmental stress and inhibitors, and an increased ability to cause infection and disease. Dermal wounds often provide an ideal environment for bacteria to exist as a community, which may have a significant effect on wound healing. — Chronic wounds are invariably polymicrobial, yet most research to date has focused on the role of specific potential pathogens in wounds (eg Pseudomonas aeruginosa) rather than the effect of interactions between different species. Only recently have microbiologists begun to address and recognise the significance and importance of an interacting community and the relationship to the disease process. It is the intention of this review to address the potential role that microbial communities and associated biofilms may play in wound healing. — The composition (chemical, biological and physical) of a surface will determine the initial microbiology that develops at that surface. In the case of a recently formed wound, different micro-organisms from endogenous and exogenous sources will contaminate the wound surface. The properties of the wound surface will predetermine which micro-organisms will attach, grow and remain components of an early biofilm. The initial colonising bacteria on any surface are referred to as the ‘pioneering’ species. Colonising bacteria will modify the habitat and create a ‘microenvironment’ that encourages the attachment and growth of secondary colonising micro-organisms. If unchallenged, and with favourable conditions, a complex community of micro-organisms is likely to develop. Although such a continuum of events has been documented in the oral cavity, microbial development and succession in wounds have not been investigated. It has been proposed that as a microbial biofilm develops the community will ultimately form a more stable ‘climax community’ [7]. As the community is able to adapt appropriately to outside perturbations the term ‘microbial homeostasis’ has been suggested to reflect stability within a ‘climax community’ [7]. — Most infections are caused by a combination of micro-organisms (polymicrobial or mixed infection) [12]. In a wound environment, individual organisms may be unable to satisfy the requirements to cause disease and subsequently combine forces to do this. Examples of this type of situation have been documented for diseases of the gum and peridontium [13] and leg ulcer infections [8]. — In many animal models numerous combinations of anaerobic and aerobic bacteria have been shown to produce levels of sepsis or disease that could not be induced by individual species [13]. Such synergy has been demonstrated in wound pathogens such as: Prevotella melaninogenicus, Porphyromonas asaccharolytica and Peptostreptococcus micros[14]; Porphyromonas asaccharolytica and Klebsiella pneumoniae[8], [15]; Escherichia coli and Bacteriodes fragilis[8], [16] and Staphylococcus aureus[8]. — Mechanisms that constitute pathogenic synergy include communal protection from phagocytosis and intracellular killing, production of essential growth factors, modification of the local environment (eg redox potential) and the protection of sensitive species by the inactivation of inhibitors. This protection has been named ‘indirect pathogenicity’ [17]. In certain situations some pathogens are found to be antibiotic sensitive but are rendered ‘resistant’ by other members of the mixed infection. Treatment in these cases would be targeted at all of the component micro-organisms in a mixed infection [17], [18]. — Opportunistic pathogens are members of the resident microflora that become pathogenic by gaining access to sites that are not normally accessible to them, or by causing changes in the local environmental conditions that favour their prevalence within a community. In addition to synergy, organisms within a wound may interact via quorum sensing.”
Again, the keys to such an incredible process of coordination among species are vesicle/exosome communications and Quorum sensing.
“Quorum sensing involves communication strategies within a mixed community of organisms that ultimately enable them to coordinate their activities, and enhance their pathogenicity and ability to cause disease.
Quorum sensing plays a major role in communications among oral bacteria and is deeply indicated in the propagation of periodontal disease and dental caries. Multiple species of bacteria live in the mouth and can interact via quorum sensing so as to produce multi-species biofilms and induce dental caries or periodontal disease. The process is highly articulated.
Biofilm-formation can lead to infected swollen gums, recession of gums and loss of teeth and accelerated tooth decay.
The 2002 publication Communication among Oral Bacteria reports: “Human oral bacteria interact with their environment by attaching to surfaces and establishing mixed-species communities. As each bacterial cell attaches, it forms a new surface to which other cells can adhere. Adherence and community development are spatiotemporal; such order requires communication. The discovery of soluble signals, such as autoinducer-2, that may be exchanged within multispecies communities to convey information between organisms has emerged as a new research direction. Direct-contact signals, such as adhesins and receptors, that elicit changes in gene expression after cell-cell contact and biofilm growth are also an active research area. – Several oral bacterial species are amenable to genetic manipulation for molecular characterization of communication both among bacteria and between bacteria and the host. A successful search for genes critical for mixed-species community organization will be accomplished only when it is conducted with mixed-species communities. — Communication is a key element in successful organizations. The bacteria on human teeth and oral mucosa have developed the means by which to communicate and thereby form successful organizations. These bacteria have coevolved with their host to establish a highly sophisticated relationship in which both pathogenic and mutualistic bacteria coexist in homeostasis. The fact that human oral bacteria are not found outside the mouth except as pathogens elsewhere in the body (51) points to the importance of this relationship. Communication among microorganisms is essential for initial colonization and subsequent biofilm formation on the enamel surfaces of teeth and requires physical contact between colonizing bacteria and between the bacteria and their host. Without retention on the tooth surface, the bacteria are swallowed with the saliva. Through retention, these bacteria can form organized, intimate, multispecies communities referred to as dental plaque. — Sequential changes in populations of bacteria associated with tooth eruption (20, 21, 102, 138) as well as with caries development (53) and periodontal disease states (109, 136) are known. Temporal changes in populations of bacteria on tooth surfaces after professional cleaning are ordered and sequential (92, 114, 115). Such sequential changes must occur through attachment and growth of different bacterial species. With the attachment of each new cell type, a nascent surface is presented for the attachment of other kinds of bacteria, resulting in a progression of nascent surfaces and concomitant changes in species diversity (79, 137). Such coordination indicates communication. In the absence of communication, these orderly changes would be random. Due to the dynamics of growth and adherence, the bacterial populations in the oral cavity are constantly changing, even during the intervals between normal daily oral hygiene treatments. It is unlikely that the various species within oral biofilms function as independent, discrete constituents; rather, these organisms function as a coordinated community that uses intra- and interspecies communication. — For the past 40 years, pure cultures of oral bacteria have been isolated from supragingival and subgingival dental plaque removed from healthy and diseased sites. The numbers and variety of bacteria obtained from many clinical conditions have been catalogued (10, 19, 72, 109, 137). Estimates of the bacterial species diversity in the oral cavity, based on both culture-dependent methods (109, 136) and culture-independent methods (83, 123), indicate about 500 species. About 415 species are estimated to be present in subgingival plaque (123), and many of these are also found in supragingival plaque. Cultured species account for about 60% of the organisms identified by molecular methods, indicating that the oral cavity is an environment where most species can be studied by routine culture methods. This is distinct from other environments where less than 1% of the clones obtained by molecular methods represent cultured species (2, 44, 54, 118). Thus, dental plaque is one of the best-described mixed-species bacterial communities. Because it is easily accessible, it is convenient to study this complex model system.”
The 2008 publicationBacterial quorum sensing: signals, circuits, and implications for biofilms and disease reported: “Communication between bacteria, belonging to the same species or to different species, is mediated through different chemical signals that are synthesized and secreted by bacteria. These signals can either be cell-density related (autoinducers) or be produced by bacteria at different stages of growth, and they allow bacteria to monitor their environment and alter gene expression to derive a competitive advantage. The properties of these signals and the response elicited by them are important in ensuring bacterial survival and propagation in natural environments (e.g., human oral cavity) where hundreds of bacterial species coexist. First, the interaction between a signal and its receptor is very specific, which underlies intraspecies communication and quorum sensing. Second, when multiple signals are synthesized by the same bacterium, the signaling circuits utilized by the different signals are coordinately regulated with distinct overall circuit architecture so as to maximize the overall response. Third, the recognition of a universal communication signal synthesized by different bacterial species (interspecies communication), as well that of signals produced by eukaryotic cells (interkingdom communication), is also integral to the formation of multispecies biofilm communities that are important in infection and disease. The focus of this review is on the principles underlying signal-mediated bacterial communication, with specific emphasis on the potential for using them in two applications-development of synthetic biology modules and circuits, and the control of biofilm formation and infection.”
Again from Interactions between Oral Bacteria:Inhibition of Streptococcus mutans Bacteriocin Production by Streptococcus gordonii: “Streptococcus mutans has been recognized as an important etiological agent in human dental caries. Some strains of S. mutans also produce bacteriocins. In this study, we sought to demonstrate that bacteriocin production by S. mutans strains GS5 and BM71 was mediated by quorum sensing, which is dependent on a competence-stimulating peptide (CSP) signaling system encoded by the com genes. We also demonstrated that interactions with some other oral streptococci interfered with S. mutans bacteriocin production both in broth and in biofilms. The inhibition of S. mutans bacteriocin production by oral bacteria was stronger in biofilms than in broth. Using transposon Tn916 mutagenesis, we identified a gene (sgc; named for Streptococcus gordonii challisin) responsible for the inhibition of S. mutans bacteriocin production by S. gordonii Challis. Interruption of the sgc gene in S. gordonii Challis resulted in attenuated inhibition of S. mutans bacteriocin production. The supernatant fluids from the sgc mutant did not inactivate the exogenous S. mutans CSP as did those from the parent strain Challis. S. gordonii Challis did not inactivate bacteriocin produced by S. mutans GS5. Because S. mutans uses quorum sensing to regulate virulence, strategies designed to interfere with these signaling systems may have broad applicability for biological control of this caries-causing organism.”
Inhibiting quorum sensing among oral bacteria may be a strategy for prevention of dental caries and periodontal disease.
The 2010 publication Antimicrobial and antibiofilm activity of quorum sensing peptides and Peptide analogues against oral biofilm bacteriareports: “Widespread antibiotic resistance is a major incentive for the investigation of novel ways to treat or prevent infections. Much effort has been put into the discovery of peptides in nature accompanied by manipulation of natural peptides to improve activity and decrease toxicity. The ever increasing knowledge about bacteria and the discovery of quorum sensing have presented itself as another mechanism to disrupt the infection process. We have shown that the natural quorum sensing (QS) peptide, competence-stimulating peptide (CSP), used by the caries causing bacteria Streptococcus mutans when used in higher than normally present concentrations can actually contribute to cell death in S. mutans. Using an analogue of this quorum sensing peptide (KBI-3221), we have shown it to be beneficial at decreasing biofilm of various Streptococcus species. This chapter looks at a number of assay methods to test the inhibitory effects of quorum sensing peptides and their analogues on the growth and biofilm formation of oral bacteria.”
Sub-gingival biofilms formed on tooth surfaces have full multispecies complexity and are responsible for gum diseases and tooth decay
This blog entry is long enough for now and, I believe, contains important messages. There is more to be said about quorum sensing and exosome communications, however. Topics I will likely cover in a future Part 3 quorum sensing blog entry include:
roles of QS in cancer and how deregulated QS may lead to tumorigenesis,
The importance of QS in organizing immune system responses, and
The important roles of QS in stem cell maintenance and differentiation.
The purpose of this blog entry is to lay out a hypothesis that could turn out to be very significant for health and longevity. The evidence cited here for this hypothesis is still initial but already compelling. We believe that this hypothesis is potentially of a breakthrough nature because of the additional insight it provides and because it can potentially lead to highly practical measures to improve the healthfulness of foods significantly. The hypothesis is original to the authors here.
As we learn more and receive reactions to this hypothesis from readers and colleagues, we expect to update this blog post to make it more complete. The current update is for August 22, 2013.
Our hypothesis is summarized in eleven points:
Most of the cells and even much of the organic structure of many fruits and vegetables remain alive right up to the point where they are destroyed by stomach acid or bacteria and are digested in our gut. Of course, as fruits and vegetables decay their cells will die.
These cells remain capable of producing the same plant-based stress responses that they produce in live plants.
In many cases for fruits and vegetables, the stress responses include plant cells producing phytochemicals such as lycopene in the case of tomatoes, pterostilbene in the case of blueberries, and allicin in the case of garlic. Such phytochemicals can be thought of as plant stress-responsive hormones.
In many instances, stress can increase the production of such health-producing phytochemicals by an order of magnitude or more.
Such phytochemicals produced as a result of plant cell stress have numerous documented health benefits for humans, and possibly also longevity benefits. Numerous entries in this blog have discussed these benefits in various contexts.
The phytochemicals address evolutionarily conserved pathways in us working via xenohormesis. That is, we respond to many plant stress-produced phytochemicals in similar ways to how the plants themselves respond.
Besides upregulated production of health-inducing phytochemicals, stresses on live fruit and vegetable cells have a second mode of health-producing action. In the process of eating plants, miRNA components are released in microvesicles which enter and show up in the circulatory systems of animals who eat them. These micro RNAs encode for multiple epigenetic impacts including health-producing ones
Therefore, stresses experienced by fruits and vegetables experienced from the point of cultivation to the point of digestion in the gut may have major impacts on the health-producing properties of such fruits and vegetables for us humans. Xenohormetic activity of a consumed fruit or vegetable cell could result from stress responses in a plant cell right up to a point where that cell is digested and dies.
Stresses on fruits or vegetables associated with cultivation, storage, transportation, industrial processing, cooking, freezing, and other forms of food preparation and even chewing and encounters with the gut bacteria might have significant impacts on either degrading or enhancing the contributions of these foods to human health. Stresses on fruits and vegetables that activate known stress responses include heat, cold, UV exposure, desiccation, cutting and slicing, exposure to bacteria, viruses, chewing, and acid exposure. Experienced stresses can be many and vary by specific fruit and vegetable. Our hypothesis is that in many cases such stresses may greatly enhance or denigrate the healthiness associated with consumption.
Such stresses are mostly open for manipulation by us, allowing us to decrease or increase them or change the timing for when they occur.
The economic consequences of our learning how to manage these stresses to benefit humans could be enormous, involving trillions of dollars.
From the blog entry Part 1: Slaying two dragons with one stone:“Xenohormesis is the concept that different species such as plants and animals have common stress signaling molecules, and that such molecules can be harvested from plants and used to increase stress adaptation pathways in animals(ref)(ref)(ref). Plants cannot run away from predators, parasites, infectious agents, hot weather, cold weather or dryness. For this reason, plants have evolved a large number of molecular stress coping pathways that are activated by compounds that are actively synthesized in response to the stressor. Some of these compounds ward off predators with bitter tasting compounds. Others ward off potential organisms that would eat the plant by synthesizing poisons that would kill the predator (i.e. natural pesticides). Others – the ones of main interest to us – strengthen the plant and allow survival in the face of the stress. Although there are many toxic compounds in this arsenal of phytochemicals, there is a large family of molecules called polyphenols that are non-toxic and appear to have great benefits in humans. Approximately 14,000 of these plant-based stress-signaler polyphenol compounds have been discovered so far in plants. They are found in the leaves, stems, flowers, seeds, fruits, nuts, and shells surrounding the nuts. These plant polyphenols appear to be xenohormetic compounds in that they also upregulate stress coping pathways in mammalian cells. These xenohormetic compounds appear to prevent aging and cancer through a large number of pathways. For this reason, their mechanism of action is multifactorial or pleiotropic. Xenohormetic compounds include resveratrol, curcumin, EGCG, isothiocyanates, secoiridoids, genistein, gallic acid, lycopene, allyl mercaptan, plumbagin, etc. Multiple plant polyphenols and their mechanisms of action have been reviewed in past entries in this blog. See for example ref, ref, ref, ref, ref, ref, ref and ref.
“A surprising number of plant molecules in our diet interact with key regulators of mammalian physiology to provide health benefits. Shown are three examples: resveratrol found in numerous plants and concentrated in red wine; curcumin from turmeric; and epigallocatechin-3-gallate (EGCG) in green tea. These compounds modulate key pathways that control inflammation, the energy status of cells, and cellular stress responses in a way that is predicted to increase health and survival of the organism. Such observations raise the question: are these biochemical interactions merely a remnant of what existed in the common ancestor of plants and animals, or is selection maintaining interactions between the molecules of plants and animals? Some interactions activate signaling pathways (arrows) whereas others inhibit them (bars). Solid arrows or bars indicate instances where there is some evidence of a direct interaction of the plant metabolite with a mammalian protein(ref)” A mechanism of xenohormesis appears to be animal gene regulation mediated by plant micro RNAs acquired through food intake.” More yet on xenohormesis can be found in that blog entry.
There is another health action of eating live fruit and vegetable cells: Beneficial miRNAs and microvesicle communications from plant cells
So far, we have discussed how stresses on fruits and vegetables, say induced in the processes of cooking or slicing, could greatly increase the content of health producing phytochemicals & that would be ingested upon eating the fruits and vegetables. Presumably, a human being could get the same substances in dietary supplement pills and capsules. However, there is an additional part of the story that goes beyond the actions of phytochemicals. And that part involves plant-based miRNAs which are likely to be communicated directly from live plant cells via secreted vesicles. It turns out that a significant fraction of our circulating in miRNAs are plant-based. Again from a recent blog entry: The 2011 publication Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNAreports: “Here, we report the surprising finding that exogenous plant miRNAs are present in the sera and tissues of various animals and that these exogenous plant miRNAs are primarily acquired orally, through food intake. MIR168a is abundant in rice and is one of the most highly enriched exogenous plant miRNAs in the sera of Chinese subjects. Functional studies in vitro and in vivo demonstrated that MIR168a could bind to the human/mouse low-density lipoprotein receptor adapter protein 1 (LDLRAP1) mRNA, inhibit LDLRAP1 expression in liver, and consequently decrease LDL removal from mouse plasma. These findings demonstrate that exogenous plant miRNAs in food can regulate the expression of target genes in mammals.” The miRNAs are present in microvesicles in the plasma where they are circulated to various cell types and serve as intercellular signaling molecules. “Our further studies demonstrated that miRNAs could be selectively packaged into MVs and actively delivered into recipient cells where the exogenous miRNAs can regulate target gene expression and recipient cell function15. Thus, secreted miRNAs can serve as a novel class of signaling molecules in mediating intercellular communication15. The novel and important functions of the secreted miRNAs were also reported by many other groups18,19,20,21. The identification of circulating miRNAs, mainly delivered by cell-secreted MVs, as stable and active signaling molecules opens a new field of research in intercellular and interorganelle signal transduction.”
The implication of microvesicle-communicated plant-based miRNAs for epigenetic regulation of human gene expression are profound. “MicroRNAs (miRNAs), a class of 19-24 nucleotide long non-coding RNAs derived from hairpin precursors, mediate the post-transcriptional silencing of an estimated 30% of protein-coding genes in mammals by pairing with complementary sites in the 3′ untranslated regions (UTRs) of target genes1,2. miRNAs have been widely shown to modulate various critical biological processes, including differentiation, apoptosis, proliferation, the immune response, and the maintenance of cell and tissue identity1,2. Dysregulation of miRNAs has been linked to cancer and other diseases3,4. Recently, we and others found that mammalian miRNAs exist stably in the sera and plasma of humans and animals5,6. – We next characterized the possible carrier of circulating miRNAs. Microvesicles (MVs) are small vesicles that are shed from almost all cell types under both normal and pathological conditions13,14. They bear surface receptors/ligands of the original cells and have the potential to selectively interact with specific target cells and mediate intercellular communication by transporting bioactive lipids, mRNA, or proteins between cells13,14. Our recent results demonstrated that MVs from human plasma are a mixture of microparticles, exosomes, and other vesicular structures and that many types of MVs in human plasma contain miRNAs15. These findings were in agreement with the recent reports by other investigators that exosomes from cultured cells served as physiological carriers of miRNAs16,17. — miRNAs are present in human and animal sera and organs. Upon investigation of the global miRNA expression profile in human serum, we found that exogenous plant miRNAs were consistently present in the serum of healthy Chinese men and women. As shown in Figure 1A and Supplementary information, Table S1, Solexa sequencing revealed ~30 known plant miRNAs in Chinese healthy donors, among which MIR156a and MIR168a showed considerable levels of expression(ref).”
Because they are packaged in microvesicles, the plant miRNAs in food can go where they will in the body. They can pass unharmed through the GI track resisting gut bacterial action and stomach acids, enter the sera and organs and pass through the blood-brain barrier.
Microvesicles, like their human cousin exosomes, are generated in live cells and released under stress conditions.
So, there appears to be two operating mechanisms of stress-induced xenohormesis connected with eating fruits and vegetables: production of beneficial phytochemicals, and release of exosomes containing beneficial miRNAs. The second mechanism requires our direct contact with live fruit and vegetable cells.
About phytochemicals and how they work
The benefits of consuming fruits and vegetables have universally been attributed to unique phytochemicals they contain: plant polyphenols involving substances like flavonoids, anthocyanins, Isoflavones, indoles, saponins, isocynates, sesquiterpene lactones, anacardic acid. etc. – along with minerals and essential micro-nutrients. These substances produce beneficial effects and activate stress-response pathways like Nrf2. This is well and good. However we are suggesting that there may be an additional and deeper level of explanation for the health-producing actions of fruits and vegetables, That level of explanation goes beyond the chemical-action theory and could provide us with insights that allows us new forms of control over the health benefits of foods we consume.
Our concept is an extension of xenohormesis, the notion that evolutionarily-conserved plant-based stress responses can be elicited in humans who eat the plants. The essence of our new idea is that the plant cells in fruits and vegetables are mostly alive before we cook, eat or digest them, and are therefore likely to be stress-responsive. Our manipulation of such stresses can enhance the healthfulness of many fruits and vegetables. Below, we cite some examples that show foods are stress-responsive to steps in cooking and preparation.
Preliminary evidence for the hypothesis: On food preparation and stress-induced impacts relating to healthfulness
Indeed, there appears to be evidence that food preparation and cooking can both positively and negatively impact human health. The following examples and quotes are drawn from the book Eating on the Wild Side – a Field Guide to Nutritious Foods.
Garlic: Allicin is a key cancer-fighting component of garlic. Allicin is created when two substances in garlic come into contact with each other: a protein fragment alliin and a heat sensitive enzyme alliinase. “in an intact clove of garlic these compounds are isolated in separate compartments. They do not commingle until you slice, press or mince the garlic.” Then the allicin-producing reaction occurs. “Israelis discovered that heating garlic shortly after crushing it or slicing it destroys the heat-sensitive enzyme that triggers the reaction. As a result no allicin is created.” Two minutes in the frying pan or 30 seconds of microwaving wipes out allicin production and ability to thin blood almost completely. The best way to get the benefits of garlic are to chop and mash it and then let it sit for 10 minutes away from heat. This creates the maximum amount of allicin. After the allicin is created, the heat-sensitive enzyme is no longer needed and the garlic may be cooked without losing the allicin and its beneficial properties. In this case, the stress producing a beneficial result is chopping or mincing. We wonder if the allicin production is an adaptive stress response, say to a garlic plant being eaten by an animal. Allicin gives garlic its strong taste and smell.
Onions: Here, we have a different but still interesting situation. An important health-producing ingredient in onions is quercetin. “Studies have shown that baking, sautéing roasting or frying onions increases their quercetin content. It is not clear to me whether this is due to a heat-induced chemical reaction or due to a hormetic heat-shock response in the onion cells. Boiling is the only cooking method that reduces quercetin content. The quercetin leaches out into the water when onions are boiled. Strong pungent onions have the most phyto-nutrients but these can be neutralized by boiling.” Here, the upgraded adaptive stress response in onions appears to be due to heat shock in response to cooking.
Corn: Canning can enhance phytonutrient content, the opposite of what is commonly assumed. “It is now clear that vitamin C provides only a fraction of the antioxidant power in most fruits and vegetables. The majority comes from phytonutrients.” (We of course now know that the phytonutrients are not themselves antioxidants but that they active production of endogenous antioxidants via the Nrf2 and possibly other pathways. Moreover, there is the recent paper to the effects that these “antioxidants” are in fact ROS-inducing pro-oxidants) “ – many phytonutrients maintain their antioxidant properties when they are heated. Some even become even more potent because the heat transforms them into more-active forms or easier to absorb. This explains why canned corn is higher in carotenoids than fresh corn.” Again, to us this explanation falls flat. It is not clear to us whether the impact on corn is due to a heat-induced chemical reaction or due to a hormetic heat-shock response in the corn cells.
Tomatoes: Tomatoes provide a particularly interesting case-in-point. A key health=producing phytochemical in tomatoes is lycopene. “Tomatoes, like a few other fruits, are better for you cooked than raw. *In fact, the longer you cook them, the more health benefits you get. They heat increases the food value in two ways. First, it breaks down the fruit’s cell walls, making their nutrients more bioavailable. Second, it twists the lycopene molecule into a new configuration that is easier to absorb. — Thirty minutes of cooking can more than double their lycopene content — .” The explanation given in the book is that the heat converts trans lycopene into cis-lycopene, the more bioavailable form. Again, it is not clear to us whether this is purely a chemical reaction or involves a tomato cell heat stress response, or perhaps both. In any event, it is yet another example of how stress in food preparation can improve the health producing-properties of a food or vegetable. Continuing from the book: the most nutritious tomatoes in the supermarket are not in the produce section – they’re in the canned goods aisle. Processed tomatoes, whether canned or cooked into a paste or sauce, are the richest known sources of lycopene. The reason is that the heat of the canning process makes the lycopene more bioavailable. — Tomato paste, the most concentrated form of processed tomato, has up to 10 times more lycopene than raw tomatoes. Tomatoes produce lycopene to protect themselves from UV rays.” (Production of lycopene is clearly a stress response.) As to the xenohormetic affect, “eating tomato paste has the same effect on us. In a German study, volunteers were divided into two groups. Half the participants made no changes in their eating habits. The other half added 3 tablespoons of tomato paste to their daily diets. When the volunteers were exposed to enough UV rays to produce a modest sunburn, the people who had been consuming tomato paste were 40% less red overall.” Tomatoes provide an excellent example of xenohormesis at work.
The 2002 study Thermal Processing Enhances the Nutritional Value of Tomatoes by Increasing Total Antioxidant Activity documents the increase in lycopene in tomatoes due to cooking them. “Processed fruits and vegetables have been long considered to have lower nutritional value than their fresh commodities due to the loss of vitamin C during processing. This research group found vitamin C in apples contributed <0.4% of total antioxidant activity, indicating most of the activity comes from the natural combination of phytochemicals. This suggests that processed fruits and vegetables may retain their antioxidant activity despite the loss of vitamin C. Here it is shown that thermal processing elevated total antioxidant activity and bioaccessible lycopene content in tomatoes and produced no significant changes in the total phenolics and total flavonoids content, although loss of vitamin C was observed. The raw tomato had 0.76 ± 0.03 μmol of vitamin C/g of tomato. After 2, 15, and 30 min of heating at 88 °C, the vitamin C content significantly dropped to 0.68 ± 0.02, 0.64 ± 0.01, and 0.54 ± 0.02 μmol of vitamin C/g of tomato, respectively (p < 0.01). The raw tomato had 2.01 ± 0.04 mg of trans-lycopene/g of tomato. After 2, 15, and 30 min of heating at 88 °C, the trans-lycopene content had increased to 3.11± 0.04, 5.45 ± 0.02, and 5.32 ± 0.05 mg of trans-lycopene/g of tomato (p < 0.01). The antioxidant activity of raw tomatoes was 4.13 ± 0.36 μmol of vitamin C equiv/g of tomato. With heat treatment at 88 °C for 2, 15, and 30 min, the total antioxidant activity significantly increased to 5.29 ± 0.26, 5.53 ± 0.24, and 6.70 ± 0.25 μmol of vitamin C equiv/g of tomato, respectively (p < 0.01). There were no significant changes in either total phenolics or total flavonoids.” This is a good example of a plant stress-response hormone (lycopene) being induced by heat stress.
Tomatoes also provide an example of where industrial processing increases food value, the kind of situation we would like to see more of. See the NYT opinionNot all industrial foods are evil.
Blueberries: Blueberries are another example where the cooked fruit offers more phyto nutrients than the raw fruit. We trust the description of what happens here and in other cases in the Eating on the Wild Side book, but not necessarily the explanations given of why it happens. “Cooked blueberries, believe it or not, have greater antioxidant levels than fresh berries. Even canned blueberries are better for you than fresh packed fruit, providing you consume the canning liquid along with the berries. The reason that cooking and canning increases their nutritional content is that the heat rearranges the structure of the phyto nutrients and also makes them more bioavailable. Many other berries respond in a similar fashion.” Again, the phytochemicals in blueberries are not themselves antioxidants; they are activators of the Nrf2 pathway in humans which turn on our endogenous antioxidant-producing genes. It is those stress-responsive phytochemicals which are increased due to heat stress in cooking and canning.
How convenient in this and several others of these examples that the heat “rearranges the structure of the phytonutrients!” We strongly suspect this is a heat-stress response in the blueberry cells mediated by heat shock proteins. It probably happens when cooking is just started before the plant cells die.
Broccoli: From a 2007 Science Daily article reporting research results, Culinary Shocker: Cooking Can Preserve, Boost Nutrient Content Of Vegetables: “In the new study, the researchers evaluated the effects of three commonly-used Italian cooking practices — boiling, steaming, and frying — on the nutritional content of carrots, zucchini and broccoli. Boiling and steaming maintained the antioxidant compounds of the vegetables, whereas frying caused a significantly higher loss of antioxidants in comparison to the water-based cooking methods, they say. For broccoli, steaming actually increased its content of glucosinolates, a group of plant compounds touted for their cancer-fighting abilities. The findings suggest that it may be possible to select a cooking method for each vegetable that can best preserve or improve its nutritional quality, the researchers say.” Here again, the so-called “antioxidant compounds” are no doubt the stress-induced phytochemicals which activate endogenous antioxidant pathways, and are not themselves actual antioxidants.
There are more such examples of where cooking stress induces health-producing phytochemical content. The 2006 publication Content of redox-active compounds (ie, antioxidants) in foods consumed in the United States provides the following table. The titles of the article and table are misleading. By “antioxidant content” the article is referring to content of beneficial polyphenols which upgrade endogenous body antioxidant responses. It was fashionable back in 2006 to call these “antioxidants” though in fact some are chemically pro-oxidants.
Effects of Processing on Antioxidant Content in Foods
Food
Type of Processing
Antioxidant Content % Compared to Non-Processed Food
Apples
Peeling
(-)33-66%
Carrots
Steaming
(+)291%
Carrots
Boiling
(+)121-159%
Cucumbers
Peeling
(-)50%
Asparagus
Steaming
(+)205%
Broccoli
Steaming
(+)122-654%
Cabbage, green
Steaming
(+)448%
Cabbage, red
Steaming
(+)270%
Green pepper
Steaming
(+)467
Red pepper
Steaming
(+)180%
Potatoes
Steaming
(+)105-242%
Tomatoes
Steaming
(+)112-164%
Spinach
Boiling
(+)84-114%
Sweet potatoes
Steaming
(+)418%
The heat-shock stress response encountered in cooking appears to be very powerful in generating healthful phytochemicals. Since plant stress responses are generally concentrated on or near the skins, another general theme is that peeling reduces beneficial polyphenol content. So, whole carrots are better nutritionally than baby peeled carrots and peeling apples or cucumbers is not a good idea.
Light exposure is another stress that can increase phytosubstance content and nutritional value in several fruits and vegetables. It is particularly effective when the light is synchronized to original plant circadian response.
An additional dimension of evidence for our hypothesis is that after harvesting, fruits and vegetables continue to produce polyphenols as driven by a daily circadian rhythm. This happens best if the fruits or vegetables are exposed to light according to the daily rhthym thay are used to. We are grateful to Dan Campagnoli, a reader of this blog, who responded in a comment below mentioning two citations.
One of these citations is the June 2013 publication Postharvest Circadian Entrainment Enhances Crop Pest Resistance and Phytochemical Cycling. “The modular design of plants enables individual plant organs to manifest autonomous functions [1] and continue aspects of metabolism, such as respiration, even after separation from the parent plant [2]. Therefore, we hypothesized that harvested vegetables and fruits may retain capacity to perceive and respond to external stimuli. For example, the fitness advantage of plant circadian clock function is recognized [3,4]; however, whether the clock continues to influence postharvest physiology is unclear. Here we demonstrate that the circadian clock of postharvest cabbage (Brassica oleracea) is entrainable by light-dark cycles and results in enhanced herbivore resistance. In addition, entrainment of Arabidopsis plants and postharvest cabbage causes cyclical accumulation of metabolites that function in plant defense; in edible crops, these metabolites also have potent anticancer properties [5]. Finally, we show that the phenomena of postharvest entrainment and enhanced herbivore resistance are widespread among diverse crops. Therefore, sustained clock entrainment of postharvest crops may be a simple mechanism to promote pest resistance and nutritional value of plant-derived food.” Entrainment here involves the synchronization of the internal rhythm of responsiveness (which occurs, say, in cabbage after harvesting) with the light-cycle external environment.
The document says this entrainment works even after harvesting in some plants to increase phytochemical content. The light stress produces protective polyphenols and facilitates the process of the fruit or vegetable ripening. For experienced cooks, there is little here of practical importance that is new. Of course, tomatoes that fall off the vine continue to ripen on the ground. And you can leave green tomatoes out of the refrigerator and out in the light where they wlll ripen and turn red – at the same t so what you want to do is good library get a book while and go to the Mexican restaurant and then go before cooking and I felt that has to be beautiful young you what were well right now what were doing ime increase their lycopene content.
The other item citation by Dan is Does your salad know what time it is? This is a recent press release on research at Rice University. “Vegetables and fruits don’t die the moment they are harvested,” said Rice biologist Janet Braam, the lead researcher on a new study this week in Current Biology. “They respond to their environment for days, and we found we could use light to coax them to make more cancer-fighting antioxidants at certain times of day.” Braam is professor and chair of Rice’s Department of Biochemistry and Cell Biology. — Braam’s team simulated day-night cycles of light and dark to control the internal clocks of fruits and vegetables, including cabbage, carrots, squash and blueberries. The research is a follow-up to her team’s award-winning 2012 study of the ways that plants use their internal circadian clocks to defend themselves from hungry insects. That study found that Arabidopsis thaliana — a widely used model organism for plant studies — begins ramping up production of insect-fighting chemicals a few hours before sunrise, the time that hungry insects begin to feed.”
The examples and data cited are sufficient to make our point with respect to cooking and light stresses. Other stresses also probably strongly impact phytochemical content and plant cell microvesicle production, such as cropping, refrigeration, supermarket preparation and even chewing, but the possible positive impacts of such stresses appear to be little studied. We believe further studies related to such stresses might reveal important new insights. Finally, there is a whole question of how live consumed fruit and vegetable cells produce stress reactions in response to stomach acids and gut biome microbial constituents – another potentially fruitful area for new research.
Consumption of fruits and vegetables could have an enormous economic impact
The Union Of Concerned Scientists has just put out a report THE $11 TRILLION REWARD arguing that the economic value of people consuming even slightly more fruits and vegetables would have incredible economic value for the society. Among the report’s findings are:
More than 127,000 deaths per year from cardiovascular diseases could be prevented, and $17 billion in annual national medical costs could be saved, if Americans increased their consumption of fruits and vegetables to meet existing dietary recommendations.
Using estimates of how much people are willing to invest in measures to reduce cardiovascular disease mortality, the present value of lives saved in the above-bulleted way would exceed $11 trillion.
And, of course, there are many other positive health impacts to consuming fruits and vegetables besides those on the cardiovascular system, implications for the reduction of cancer, diabetes, neurodegenerative diseases and wound healing. So the ultimate health and economic benefits would be much greater yet. We are here suggesting the possibility that even greater benefits could be realized through careful management of the stresses experienced by fruits and vegetables before they are digested.
Conclusions
Our concept is that cells in fruits and vegetables often remain alive and active until cooked or digested, and can generate stress responses at every stage along the way from cultivation to digestion. These stresses could be associated with separation from the original soil and plant biome, physical picking, packing, cleaning and storage, movement during transportation, cold shock due to refrigeration or freezing, food preparation steps as above, cooking, chewing, and encounters with our gut biome. These stressors could include physical maceration and cutting, heat, cold, UV exposure and xenobiotic exposure. Stress responses in plant cells can include enhanced production of beneficial phytochemicals and production and dissemination of microvesicles containing plant-based miRNAs. Both can have major impacts on human gene expression. Therefore, we suggest that stresses encountered along the way by these foods from field to gut can make major contributions to enhancing or degrading the ultimate xenohormetic responses of humans. Finally, it is likely possible that we can manipulate and control these stresses so as to produce optimal health impacts. These ideas suggest new areas for possibly important research.
That cooking and light exposure can increase the nutritional values of several fruits and vegetables has been known for some time and have been regarded as partial paradoxes. We believe that the live-cell stress hypothesis laid out in this blog entry explains why these happen.
What we are suggesting is that steps in the food delivery and preparation chain for humans pre-condition the healthfulness of fruits and vegetables in specific ways due to live plant cell stress responses. Food scientists have largely been oblivious to the molecular mechanisms of such stress preconditioning on foods. And they understand the net results of such preconditioniong only in special cases, such as related in the examples given above connected with food preparation and cooking. Further research could possibly assist us in improving such pre-conditioning, giving us fruits and vegetables which produce enhanced health benefits. The health consequences and economic benefits of doing so could be very large.
I believe the the message of the previous blog entry Health through stressing fruits and vegetables – the Xenohormetic Live Food Hypothesis by Vince and Melody is worthy of acclaim. As so frequently happens, it fueled my “why gene mutation” and set me thinking and further researching. Following are some observations that expand on the actions and mechanisms of plant polyphenols such as are in fruits and vegetables. I also cite research on additional stressors not covered in the original blog entry that can affect fruits and vegetables including physical wounding, ethylene gas, root hypoxia, osmotic stress, cold stress, drought and pathogen attack. I believe these can apply both pre and post harvesting. I discuss how polyphenols are involved in causing leaves to become colored and die in the Fall – a form of apoptosis. I also mention some personal reactions – the interest generated in me by some of these points.
1. Polyphenol Synergism: 1 + 1 = more than 2
Plant polyphenols have synergistic effects, a fact worth emphasizing. There are many practical implications of this. The most important is that for health we need multiple polyphenols and not just one. This is why a variety of fruits and vegetables are important. Here is some evidence for synergism:
a. Flavonoid mixtures are more effective than single flavonoids in their antibacterial effects. For instance:
quercetin + quercitrin are better than either flavonoid alone
quercetin + morin are better than either flavonoid alone
quercetin + rutin are much better than either flavonoid alone
Rutin alone has NO antibacterial effect, but when combined with quercetin and morin, there is a greater antibacterial effect.
b. Flavonoid mixtures are more effective than single flavonols in their anticancer effects.
For instance, quercetin + kaempferol reduces the risk of cancer more than the predicted additive effects of the two polyphenols in in vitro testing of GI cancer cells and breast cancer cells.
2. Wounding lettuce induces polyphenol synthesis:further evidence that polyphenols are stress hormones
This is right in line with what was said in the previous blog with respect to garlic. The publication Effect of Intensity and Duration of Heat-shock Treatments on Wound-induced Phenolic Metabolism in Iceberg Lettuceseemed mainly focused on going over the effects of extreme heat on lettuce, but what interested me was that just simply excising the midrib section of lettuce leaves induces polyphenol synthesis. As expected, too much heat stopped this from occurring. (i.e. greater than 40 C). From 20-40 C, the increase in polyphenol synthesis from wounding was maintained, but above 40 C, the amount of polyphenol synthesis from wounding dramatically was reduced. Again, this suggests that polyphenol synthesis is a hermetic stress response for lettuce, the stress in this case being wounding via the removal of lettuce leaves.
3. Ethylene is the plant hormone that induces polyphenol synthesis in response to wounding
The ancient Egyptians used to gash figs in order to stimulate ripening. They had figured out that this wounding process would make them ripen faster. The Chinese would burn incense in closed rooms to enhance the ripening of pears. In 1901, a Russian scientist named Dimitry Neljubow showed that the compound responsible for this ripening was ethylene. However, it was not known until 1934 that plants actively synthesized ethylene and that this was not just a byproduct of plant metabolism (this sounds much like the free radical story, nitric oxide story,carbon monoxide story, and hydrogen sulfide story). Of interest is the fact that the ethylene is produced by plants in very small amounts (I do not know exactly how much, but I am sure it is much like ROS, RNS, CO, H2S, etc.)
References:
Neljubov D. (1901). “Uber die horizontale Nutation der Stengel von Pisum sativum und einiger anderen Pflanzen”. Beih Bot Zentralbl10: 128–139.
Doubt, Sarah L. (1917). “The Response of Plants to Illuminating Gas”. Botanical Gazette63 (3): 209–224.
Gane R. (1934). “Production of ethylene by some fruits”. Nature
Vince told me an interesting story related to banana boats. Bananas release ethylene gas while ripening, and the gas itself promotes ripening. There was a time when entire boatloads of bananas being shipped from Central America arrived in New York overripe or mushy due to the ethylene So, refrigeration was introduced early in banana boats and. Also, maintainance of fresh air circulation in banana boats is now recognized as important. Control of ethylene is what makes possible our access to bananas. “Without refrigerated transport, shiploads of fruit frequently arrived at best overripened, at worst in a downright rotten state, making the mass marketing of bananas impossible. With the preservative power of refrigeration and the speed of steam-powered engines, however, bananas could be shipped in enormous volumes. [...] In a matter of decades, refrigerated vessels had helped turn what in the 1890s was an exotic curiosity into a mass-market product, paving the way for a massive and highly lucrative trans-American trade(from Moveable Feasts).”
Ethylene production can also significantly reduce the shelf life of fresh flowers, potted plants and many fruits and vegetables. This plant hormone has many effects and appears to play a major role in maturation.
4. Many Environmental stressors induce ethylene synthesis by plants
Once I found out that ethylene was actively synthesized by plants, that ethylene synthesis was stimulated byplant wounding, and that ethylene induced plant polyphenol synthesis, I thought I had everything figured out, but this just raised more questions that it answered. Specifically, most plants are not wounded. Why would a pathway for ethylene synthesis occur in nature? There had to be more than just a “wounding response” that would induce selection pressure for such a biochemical pathway to evolve. Well, here is what I found out:
Many environmental cues induce ethylene synthesis in plants:
“An ethylene signal transduction pathway. Ethylene permeates the membrane and binds to a receptor on the endoplasmic reticulum. The receptor releases the repressed EIN2. This then activates a signal transduction pathway which activates a regulatory genes that eventually trigger an Ethylene response. The activated DNA is transcribed into mRNA which is then translated into a functional enzyme that is used for ethylene biosynthesis(ref)”
5. Plants respond to cold stress as well as heat stress by upgrading polyphenol production
The 2000 publication Resistance to cold and heat stress: accumulation of phenolic compounds in tomato and watermelon plantsreports: “Tomato plants, Lycopersicon esculentum L. cv. Tmknvf2, and watermelon plants, Citrullus lanatus [Thomb.] Mansf. cv. Dulce maravilla, were grown for 30 days at different temperatures (15, 25 and 35°C). We analysed soluble phenolics, enzymatic activities(phenylalanine ammonia-lyase, polyphenol oxidase and peroxidase), and dry weight. The impact of the three temperatures was different in tomato and watermelon. Our results indicate that heat stress in tomato plants occurred at 35°C, while chilling stress occurred in watermelon plants at 15°C. Thermal stress in both plants caused: (1) decreased shoot weight; (2) accumulation of soluble phenolics; (3) highest phenylalanine ammonia-lyase activity; and (4) lowest peroxidase and polyphenol oxidase activity. These results indicate that thermal stress induces the accumulation of phenolics in the plant by activating their biosynthesis as well as inhibiting their oxidation. This could be considered an acclimation mechanism of the plant against thermal stress.”
6. ROOT HYPOXIA!
Now I am getting really excited! I can’t sleep I am so excited! This is my favorite stressor at the moment! Here is how it goes:
1. With flooding, roots suffer from a lack of oxygen (anoxia).
2. Hypoxia/anoxia induces the synthesis of 1-aminocyclopropane-1-carboxylic acid (ACC) in the flooded roots.
3. The roots synthesize ACC (1-Aminocyclopropane-1-carboxylic acid) from S-adenosyl-L-methionine (SAM), our old methyl donor friend.
4. The roots synthesize SAM from the amino acid methionine.
5. ACC is transported upwards in the plant and then oxidized in the leaves
6. An end result is production of ethylene.
Conclusion: Ethylene is the “upstream” stress hormone that induces polyphenol synthesis in plants. Its production is downstream of some stresses like root hypoxia. To use a human biochemical analogy, Ethylene is the plant “ACTH” for the induction of polyphenols, which are the plant’s “cortisol.” The Ethylene is “upstream” and the polyphenols are “downstream” compounds synthesized in response to stresses.
It seems fairly clear that if you pull a plant up by its roots, like a carrot or turnip or onion, root hypoxia can also happen.
Now this raises even more questions. For example, how is ethylene synthesized?
7. How Ethylene is Synthesized: The Yang Cycle
This is really getting interesting! The synthesis pathway for ethylene is called the Yang Cycle. After studying it, however, I think a better term would be the “SAM cycle” (see diagram below)
As you can see, the Yang cycle is really just a “spin off” from methionine synthesis. However, to synthesize ethylene, you must create the intermediate S-adenosyl-L-methionine (SAM) and then ACC. (SAM is one of the best epigenetic drugs out there – a methyl donor for DNA cytosine Methylation) Then ACC is converted to ethylene in the presence of oxygen.
Why all this biochemistry? Well the diagram above misses the entire point!
THE ROOTS DIDN’T HAVE ENOUGH OXYGEN – THAT IS WHY THEY SYNTHESIZED ACC THE LEAVES HAD ENOUGH OXYGEN, SO THEY COULD COMPLETE THE ETHYLENE “HORMONE SYNTHESIS” PATHWAY.
Summary:
In summary, root hypoxia during flooding induces the conversion of methionine to SAM, which is then converted to ACC. ACC migrates from the roots to the leaves, which do have enough oxygen, where it is converted to ethylene in the presence of oxygen. Why do leaves have enough oxygen? THEY SYNTHESIZE OXYGEN, STUPID! In other words, because plant
leaves can synthesize oxygen from carbon dioxide in chlorophylls by photosynthesis, the methionine-SAM-ACC-ethylene pathway is a way for hypoxic roots to tell the entire plant that it is time to mature and then die!Ethylene is the stress hormone that does all of that.
8. Anthocyanins are the polyphenols responsible for the fall colors of leaves
Once I found out that root hypoxia causes ACC synthesis in the roots, which migrated to the leaves to induce ethylene synthesis, which in turn induced polyphenol synthesis, I asked myself “why”? (again my Why Gene has reared it’s ugly head). Here is what I started to think about:
In many countries, there is not only a spring rain, but an autumn rain. Plants exposed to flooding need to cope with this “water stress”. Plants synthesize anthocyanins in response to this stress.
It appears that anthocyanins (once thought to just be UV sunscreens for plants) are osmotic regulators of plants. Specifically, anthocyanins can decrease leave osmotic potential.
There we go! Another “proof” that the polyphenol pathway is a stress response in plants.
But why turn into all of those fall colors and then die? Well it is now obvious – polyphenols also induce plant cell senescence.
Read on, unless you are scared!
9. Ethylene induces leaf senescence and xylem senesence (i.e. time to get rid of the leaves for the winter). Perhaps this is why polyphenols induce ROS
Xylem tissue is water and nutrient transporting tissue in vascular plants.
Here is a list of some of the things that ethylene does in plants (I eliminated the ones that were too confusing)
Summary: ethylene and the anthocyanin synthesis that the ethylene induces is a way of ending the life cycle for leaves. It is a way of saying….time to die.
What relevance does this have for us? A lot! Anthocyanins do NOT induce cellular senescence in humans, of course. But we know that high levels of many of these polyphenols induce ROS, and that this is the mechanism by which they kill cancer. Although I have not yet validated this in the plant literature (I ran out of time), my theory now is that high levels of anthocyanins produced in the Fall as a result of heat stress, water stress, etc. induce ROS production in plant leaves, which then make them die and fall off the tree. If so, this explains why polyphenols induce mammalian ARE genes and cell survival, but can also induce ROS and apoptosis in cancer. In other words, the polyphenols do the same thing to cancer cells as they do to leaves in the fall…..they kill them. I will likely research this further and create another blog entry on the topic
Conclusion:Polyphenols are pleiotropic compounds synthesized in plants in response to multiple plant stressors. Thus:
Root flooding induces hypoxia, which induces ACC synthesis from methionine via SAM. ACC can migrate to the leaves where chlorophyl can synthesize oxygen to complete the process of producing ethylene. Ethylene is actively synthesized by plants to induce polyphenol synthesis.
Polyphenols such as anthocyanins are also “osmotic stress hormones” that help them cope with with water stress.
Polyphenols are implicated in many other stress coping pathways that can also be used to induce deciduous changes with the fall. Specifically, they can induce leaf senescence and make the leaves fall off. The bright colors seen in fall leaves are all due to anthocyanins, which induce leaf senescence and death.
The mechanism by which they induce senescence is unclear, but I suspect it is a ROS-mediated induction of cellular senescence. This is why polyphenols can induce cancer cell death…..i.e. I strongly suspect it is the same as inducing leaf death.
10. Several transcription factors involved in plant polyphenol synthesis have been identified and studied
These include AtMYB12, VvMYBF12, VvMYBPA1, VvMYB5b, 2R3MBY and DkMyB4 With this knowledge, new means are being suggested for significantly increasing the polyphenol content in foods. These means are likely to be of interesting for post-harvesting interventions as well as pre-harvesting ones.
AtMYB12
The 2008 publication AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: expression in fruit results in very high levels of both types of polyphenol reports “Plant polyphenolics exhibit a broad spectrum of health-promoting effects when consumed as part of the diet, and there is considerable interest in enhancing the levels of these bioactive molecules in plants used as foods. AtMYB12 was originally identified as a flavonol-specific transcriptional activator in Arabidopsis thaliana, and this has been confirmed by ectopic expression in tobacco. AtMYB12 is able to induce the expression of additional target genes in tobacco, leading to the accumulation of very high levels of flavonols. When expressed in a tissue-specific manner in tomato, AtMYB12 activates the caffeoyl quinic acid biosynthetic pathway, in addition to the flavonol biosynthetic pathway, an activity which probably mirrors that of the orthologous MYB12-like protein in tomato. As a result of its broad specificity for transcriptional activation in tomato, AtMYB12 can be used to produce fruit with extremely high levels of multiple polyphenolic anti-oxidants. Our data indicate that transcription factors may have different specificities for target genes in different plants, which is of significance when designing strategies to improve metabolite accumulation and the anti-oxidant capacity of foods.”
3. VvMYBF1
There are some innovative scientists in Germany who have identified the transcription factor in grapes that controls polyphenol synthesis in grapes. This is part of the R2R3-MYB family and has a specific domain in it called the SG7 domain. It is unclear to me if this transcription factor is “upstream” or “downstream” from the two transcription factors listed below. I assume the transcribed polyphenols would include resveratrol, but this was not specifically studied.
A group of scientists in Australia and in France have identified the transcription factors found in grapevines that regulates the synthesis of proanthocyanidins. These are precursors in the flavanoid synthesis pathway. They have not tried over-expressing these transcription factors, but this is good news for those of us who like protoanthocyanidins. These are the molecules found in chocolate that are so good for us.
References: Publications related to the molecular mechanisms of the above and other modes of plant polyphenol production include:
11. Other Interesting Questions that May be Practical
A. Would Hot house maturation of live fruit and vegetable plants with stress allow for even greater stress responses to occur in the fruits and vegetables?
We live in the city and have practically no front yard. Our back yard is a steep bank that is untillable unless you want to roll down a 100 foot bank. For this reason, I am in the process of creating a rooftop garden. The major problem with our roof top is that it gets so darn hot up there. I initially thought this would be a real problem, but now after reading the Xenohormetic Live Food Hypothesis blog entry, I am thinking this may be a great “problem” to have! Here are some of my ideas:
1. Cultivating tomatoes with 35 C (95 F) conditions induces polyphenol synthesis and prevents polyphenol oxidation
Here is an article from Granada in 2000 that shows how growing tomatoes at 35 C induces more polyphenols than growng them at 15 C or 25 C. This was a two-fold effect in enzyme production of some of the metabolites of polyphenols. (i.e. a 100% increase). This is no surprise, since it appears that polyphenols are “stress coping molecules.” According to this article:
a) thermal stress induces polyphenol biosynthesis and
b) thermal stress inhibits the oxidation of polyphenols.
The plants did not grow very well at this temperature, however. This is why I would like to grow them at more tolerable temperatures, then “stress them” when the tomatoes have fully formed.
Comments: My study of incubating blood under hyperthermic conditions and also under hypoxic conditions has led me to a greater understanding of the heat shock response elements and the hypoxic response element genes in humans.
Whereas 30 minutes of heat exposure (i.e. a long hot tub bath) does seem to activate existing heat shock proteins, meaningful changes in gene expression secondary to the nuclear translocation of the heat-induced timerization of HSF1, HSF2, HSF3, and HSF4 take several hours to occur. Peak increases in gene expression many times do not occur for 12-24 hours. For this reason, I would think that a similar effect might be seen when plants are exposed to 110-120 degree F temperatures. 30 minutes of steaming would be good, but 12-24 hours of “hot as Hell” conditions may be even better. I would like to experiment with this. The best place to do this would be on my roof, where we could easily induce these conditions in a solar reflector heated hot house (i.e. green house).
B. Would Inducing root hypoxia with root flooding be good for polyphenol content in vegetables?
This may be bad if done too much and continuously, but intermittent flooding would probably be a good thing.
C. Would cultivating tea plants with excess ethylene produce more EGCG?
I am not sure that we can make generalizations about all polyphenol synthesis pathways. I am very interested in producing some high ECGC tea leaves. This is just a thought, since I think that it takes several days to really induce synthesis via such pathways.
I expect to publish my next blog entry on this topic soon.
This is a different kind of blog entry than the usual ones, not about the hard sciences involved in aging. Instead, it is about about a practical initiative that is related to the societal developments that are co-evolving with the biological ones with respect to aging. The three previous blog entries on The evolving narrative and social transformation of agingdetail the background to the initiative of concern here, particularly Part 3: major initiatives and activities that are reflecting and driving the changes in narratives of aging.
The original version of this post stated “The initiative described here is an entry in a competitive contest sponsored by the TEDx-Fulbright Social Innovation Challenge. The Challenge is a contest to determine who participates in giving a TED talk that will be telecast internationally describing a project that represents a major social innovation. Vince is among eight semi finalists, of which four will be offering Ted talks as well as access to key people who might assist in the funding of the project described.” Further, we invited you to vote for the project on a TEDxFulbright page. We are pleased to announce that we won the voting contest, receiving the most votes and therefore became a finalist in the challenge competition. Thank you for your votes that led to our win. Our team went to Washington DC and Vince indeed gave a short TEDx talk advocating our initiative as part of a day-long TEDxFulbright event on April 5, 2014 Though we were a finalist in the Social Innovation Challenge, we did not win the first prize. That went to a different initiative that is already operating and demonstrating clear social benefits, However, we believe that our participation as a finalist in that event engendered significant interest in SEQUEL, and resulted in personal contacts that will most likely advance the cause of the SEQUEL social initiative.. Again, thank you for your votes.
The initiative we proposed in the TEDx talk was more focused than that described here, addressed to healthy and energetic retiring baby boomers who have had a dream of launching an entrepreneurial venture but not quite knowing how to proceed. The essence of the approach is to connect them with successful younger-generation people with successful entrepreneurial experience and knowledge of how to raise venture capital.
The following description of our proposed initiative is structured as was requested by the Ted Fulbright people. First, a short video submitted related to the initiative:
Please describe the social problem that you are trying to solve.
People are living a lot longer and retirement is becoming a late-midlife, rather than an end-of-life event. The issue at stake here is what are these retiring people going to do with the rest of their lives? Retire classically and essentially drop out of the social system of productivity and possibly be a drag on society? Or continue to work or otherwise contribute socially and economically and achieve fulfillment in the process?
When Social Security was implemented in 1935 life expectancy was 64 years and the retirement age was set at 65 years. Now, life expectancy is 80 but our official retirement ages have budged little. As a consequence more and more people confronting compulsory retirement are healthy, highly functional and capable of working or otherwise contributing significantly to society for many more years. Some look for another job, do part-time work, adopt a new career, do volunteer work, become family caregivers, or move to fulfill a long-standing personal dream. Others seeing no other option may simply accept retirement, effectively resigning from social participation.
Quality of decisions made at this point in life is an immensely important issue, not only for individuals figuring out what to do with the next 20 or 30 years of their lives but for nations and the world in general..There are 76 million “Baby Boomers” in the US now reaching retirement age at a rate of about 3 million a year. On a worldwide level there are more than a billion people coming to retirement age in the coming decade. From an economic perspective, the question is whether these elders will be vital contributors to society or whether they will be an economic burden that could possibly become unsustainable. The issues are utilization of human capital and trillions of dollars of either added or foregone economic productivity.
Yet, retirement or change of career beyond the age 60 can be a major life transition – posing challenges that are unique to every individual and fraught with uncertainties. The challenges include overcoming interior psychological barriers and dealing with deeply-entrenched social perceptions about the incompetency of older people. They include self-assessment of continuing health and mental competency, financial situation, job and employment skills, professional status and reputation, educational level, family situation, entrepreneurial bent, need to care for sick parents, and dreams of “What I would really like to do.”
National and local resource organizations have started mobilizing to address this challenge and have created many interesting initiatives. Many of these initiatives are detailed in our blog entry on Major initiatives and activities that are reflecting and driving the changes in narratives of aging. . However, our sense is that the response so far is nowhere up to the challenge. We think most retirees do not utilize resources that could be available to them. Recent study indicates that only 16% of retirees in the US continue doing paid work although many more say they wanted to do that. Dealing with the psychological readjustments necessary to go for a new career late in life and knowing about available resources, and evaluating and using them to facilitate a real career transition are daunting tasks. Many retiring people do not know how even to start thinking about the situation.
Please describe your solution to the problem
Ultimately, we see the need for an integrated service which we have named SEQUEL. The SEQEL service is organized around the event of retirement or career change late in life. It involves personal interaction of an older person in career transition with a volunteer or paid retirement coach. It involves live and cyber networking with accessible employment counseling and placement services, job opportunity data bases, health and vitality evaluation and maintenance services, personal life planning experts, financial counseling services, educational counseling services for seniors, services fostering entrepreneurial startups, other individuals facing comparable issues, and other community based resource individuals and organizations. SEQUEL helps retiring people to know about and connect with relevant community resources, including government and private service agencies, health clubs, churches and synagogues, senior centers, libraries, health agencies a wide range of professionals, and peer group individuals facing similar issues. The technology involved includes a number of web tools. Some of these are for personal competency and situation self- assessments. Other tools will facilitate linking to and drawing upon community resources. The service will offer data bases of local resources, web “mashups,” and social and geolocation-based networking websites that facilitate easy networking to such resources, and proactive live community networking among resource groups and individuals. Another expected benefit of the innovation will be closer community integration of retirement support services.
However, such a comprehensive SEQUEL service must be created in the context of what exists and ultimately the brunt of serving the world’s retirement seniors must be met by nationwide and community networks operated by others. There already are a number of organizations offering a variety of interesting programs to assist people through the retirement transition. Some of the resource organizations are national, some are local, some are faith or spirituality-based, some have a psychological focus, some are focused on health issues, some are offered by individual counselors, some have large organizations like the AARP behind them. Others may be based in a local church or senior center. Together, these organizations suggest numerous positive approaches including psychological counseling, personal emotional support, self-assessment of personal resources, and health assessment. However, our impression however is that many or most retiring people are unaware of and under-utilize these resources and essentially muddle through the retirement transition as best they can.
There seems to be no end to good ideas and positive approaches. Yet our perception is that they are little used. We believe pulling together these ideas and coordinating approaches into an effective highly person-centered and community based service is needed. Such a service would combine live counseling and networking with new forms of cyber networking and empowerment. And this service has to be one that can be rolled out on a national and eventually an international basis.
We do not know how best to do that right now; we don’t think anybody does. What we are proposing is a proactive and mutual discovery process that we would engage in with many other actors already on the scene to arrive at the service mixtures that are needed on a local level. The central components of tuning the solution involves 1. Bringing the importance and nature of the problem too much greater public awareness. 2. Working with other organizations already constituted to address the problem, forming multilevel partnerships to establish how a much more effective and coordinated approach might be created, and 3. Conducting pilot programs to learn how best to create and operate the comprehensive SEQUEL service we have identified.
SEQUEL must achieve its objectives via networking with others and starting with forms of organization that already exist. For example, networking among retirement-related resource organizations is already being initiated by a local branch of ENCORE in the Boston area.
Please explain why your solutions is innovative and/or unique
In general, we see the innovation of SEQUEL to be in our approach to the problem of late-life career transition issue rather than a specific solution to the problem. We believe a solution or solutions can emerge from the actions we are proposing.
The uniqueness of we are proposing is the declaration that there is an extremely important issue that needs addressing by society, that despite many well intentioned and well-designed efforts by sizeable players, these needs are not generally being well met now, that a great deal of public awareness needs to be focused on the issue, and that working with the key stakeholder organizations we can contribute to arriving at better and more-integrated solutions that exist now. We see SEQUEL as an initially a small effort that might play a very important catalytic role in mobilizing a more effective configuration among the personal retirement related services and initiatives that are already out there.
Please describe the short-term and long-term size and scale of your social impact
In the long term there is a possibility that we could help catalyze and foster a major transformation in the perceptions of and about older people and late life transitions they undertake. The scale implications are immense since the lives of billions of people could be affected and the economic consequences could be vast – in the trillions of dollars and profoundly affecting the economies of nations.
For the shorter term, our activities that should result in results that are modest but still of importance: an increasing public consciousness of the issues connected with late-midlife career transitions. Moreover, an expected impact will be bringing enhanced awareness to other service-providing organizations of best practices for effectively structuring and coordinating their activities. And, since learning must be hands-on, we expect to work with a lot of seniors in the positive shaping of their career transitions.
Please describe your implementation plan, including past achievements, pilot project, and accomplishments. Including dates would be preferable
Past personal achievements of Vince relevant to SEQUEL include the experience of having personally gone through a major career transition late in life, at age 77 seven years ago, becoming a well-known scientists in the area of longevity research, having established a major publication vehicle related to the science connected with healthy aging, and having researched and published a number of articles related to longer life spans and their social implications. We have been meeting as a team for several months now and are committed to facilitating the social transformations expected in this century related to longer and healthier life spans. All of us bring strong commitment and passion.
The pilot project we envision will have three major components:
1. Public communications: We have already been engaging in significant public communications via online print, and our TED Fulbright talk will be another kickoff activity. We are in communication with film and documentary makers and hope to secure funding for productions. Also we will be seeking public exposure in the print and TV entertainment media. Among the activities we’re planning is a contest asking people to submit YouTube videos related to how they have successfully achieved career changes late in life. We expect to have all these activities well underway within the first year of operation of SEQUEL.
2. Stakeholder communications and coordination will start out with meetings with leaders of the main organizations currently involved on a national scale (AARP Life Reimagined Institute, the Life Planning Network, and ENCORE). We hope to meet with both national leadership of these organizations and local chapter working representatives and obtain detailed perceptions of what is working and what is not working, what are the needs for expansion, and what are the unique needs that SEQEL can play in the overall process. We expect to scale our activities so that they contribute to existing initiatives and do not attempt to duplicate them. We expect to convene or participate in workshop conferences involving representatives of these organizations within the first year. Emphasis there will be on sharing experience in empowering seniors in late midlife career transitions and retirement and exploring how services of the various organizations can be better coordinated, integrated, and delivered on the local level. Our role will be as a proactive advocate for constant service improvement, research and development, adopting new technologies, and sharing of experience.
3. Our pilot project: we expect to launch a SEQUEL pilot program on a highly local basis, say in a moderate sized town in the Boston area like Somerville, and will seek to partner with the city government and other key local and national-linked stakeholder resources. The program should be in place after 18 months. The exact form of this program will depend on the community, the partners we will be able to generate, and are learning from stakeholder communications. Our major objective will be learning about what works on a community level and identifying service models that can be rolled out elsewhere. Targets are: Launch pilot project: 1 Sept 2014; pilot project fully operational 1 April 2016. Program evaluated continuously, testing of different revenue models. First formal evaluation 1 June 2017. Rollout to other locations to follow based on learning from pilot program. An alternative option will be to participate in such a pilot program with these characteristics initiated by others, should our exploration with other actors indicate that this is the best way we can contribute
Please provide your business plan. Please explain how you plan to make your solution sustainable over the long-term including operational expenses, revenue generation, investments, etc. We understand that getting a project off the ground will cost money, but please demonstrate a plan to achieve long-term sustainability. Including dates would be preferable
How the following plans play out will depend very much on the initial rounds of stakeholder communications and communications with other organizations planning similar initiatives. We do not wish to reinvent the wheel. We do wish to see highly effective vehicles on the road.
TARGET AUDIENCE/MARKET/CUSTOMER – Seniors and others who are facing retirement or work termination or who wish to develop new career opportunities late in life. Particularly, individuals who might need to base their future work or social participation on multiple personal criteria including health and financial status, skill sets, social expectations, local resources, personal networks, and dreams of what they would like to do
STARTUP FUNDING – Foundation funding with a grant of $250,000 that should support the operation of SEQUEL through the pilot phase in the first two years. May be augmented by grants for specific local initiatives.
BUSINESS AND REVENUE MODEL
The SEQUEL service is not-for-profit based. The revenue model is a blend of individual membership fees, charges to service providers and member fees for web services and workshop and training programs. Also, royalties from service providers who reach their target audiences via SEQUEL. Possibly, advertising from web-based media.
The products and services include career counseling, access to web-based services and databases, personal guides and mentors, cyber and live workshops, task-oriented social networking websites and private web and live conference groups.
We target full economic self-sustainabiity by month 24 of full operation. By year 5 if successful, projected program has 20,000 members in multiple communities and budget of approximately $3 million and is on steep growth path. Evaluation of social impact will be based on membership growth, participation and satisfaction, and on economic sustainability
Please describe your idea/organization’s status. No preference will be given to this response.
It is an unofficial organization of individuals that have been meeting for five months now, seeking to clarify its mission and not-for-profit incorporation.
Please provide a paragraph bio for each of your team members, including their positions if applicable. Please include relevant past experiences, awards, and accomplishments that would provide further validity towards the team’s ability to succeed. Please also provide their full name, affiliations, email address, and phone number for each team member at the beginning of each of their paragraph bios.
The members of the team have been meeting regularly for five months now and are in the process of forming a Massachusetts Not-for-Profit corporation.
“Being a follower, connoisseur, and interpreter of longevity research is my latest career. I have been at this part-time for well over a decade, and in 2007 this became my mainline activity. In earlier reincarnations of my career. I was Founding Dean of a graduate school and a University Professor at the State University of New York, a senior consultant working in a variety of fields at Arthur D. Little, Inc., Chief Scientist and C00 of Mirror Systems, a software company, and an international Internet consultant. I got off the ground with one of the earliest PhD’s from Harvard in a field later to become known as computer science. To get through I had to qualify myself in hard sciences, so my studies focused heavily on quantum physics. In various ways I contributed to the Computer Revolution starting in the 1950s and the Internet Revolution starting in the late 1980s. I am now engaged in doing the same for The Longevity Revolution. I have published something like 200 books and papers as well as over 440 substantive.entries in my blog, and have enjoyed various periods of notoriety. If you do a Google search on Vincent E. Giuliano, most if not all of the entries on the first pages that come up will be ones relating to me. Relevant to the proposed project, 1. I have the experience of launching launched into a new career at the age of 77 which has been highly successful, 2. My main vehicle has been researching and publishing substantial articles in my blog www.agingsciences.com. This blog is mainly concern with the hard sciences associated with aging and I have also written extensively on the societal aspects of aging. The blog has a following of 27,000 registered subscribers, 50% of which are international, and on a typical day is accessed by 3000 – 4000 readers. And 3. I have researched societal issues associated with aging and have written about them extensively.
Chen Hua, candidate in the MPA program at the Kennedy School at Harvard, chenhua_chen@hks14.harvard
Chen’s academic program focuses on social innovation. He holds an MBA from Auckland University. With cultural and business experience in Mainland China, New Zeeland and the US, he has enjoyed successful careers as a banker, a corporate executive, and a startup entrepreneur. He is now centrally concerned with social innovation and entrepreneurship associated with the shifts in aging of central concern here. He is focusing his personal career on this, and played a central role in initiating this project. He is a strong advocate of the notion of starting a new life after retirement and promoting awareness that can facilitate a long healthy, productive and disease-free life. He lectures on longevity related topics.
Melody Winnig President Vivace Associates, Research Associate at Agingsciences.com
Melody has a Master’s Degree in Information Science from the State University of New York (SUNYAB). She has worked in the field of public relations, as a consultant in development and Internet in the US, Egypt and Latin America, and in art communications. She has in recent years focused on the science and roles of plant-based substances for creating personal and public health. She has studied lifestyle patterns of centenarians and has also focused on shifting narratives of aging associated with the first wave of baby boomers. Working closely with Dr. Giuliano, she is familiar with many of the key frontiers of science impacting on aging. And she keeps up day-to-day with basic relevant research findings and communicates frequently with longevity scientists and health practitioners about these.
Michael Giuliano Planner at Agingsciences.com and Manager for Vivace Associates
Michaelhas a Masters Degree in Economic Development and Tourism Management from Boston University. He brings a business background, having worked in real estate as a broker/agent for the last 10 years relating to diverse client populations. An important contributor to the SEQUEL team, he has experience in communicating with and organizing project stakeholders, operational and project management. For the last three years, Michael has worked with the team doing research and analysis. He applies his skills in leadership, research and analysis to the role of Operations and Project Manager on our team.
Members of our team recently have just generated a three-part series of blog articles directly relevant to the project being proposed. The evolving narrative and social transformation of aging. Use these links for access. In Part 1 we started by laying out main issues confronting the world that are associated with increasingly aging populations. We do this by reviewing landmark UN and US government studies offering a global perspective on population aging. In Part 2 we list a number of the contrasting narratives related to aging which we believe to be in flux. In Part 3, we identify some of the major organizations, initiatives and activities that are reflecting and driving the changes in narratives.
By Vince Giuliano with inputs from James P Watson, Victor, and Melody Winnig
Many of our most basic biological processes including gene activation are affected by circadian and other body and cell-level clock rhythms. In January 2012, Victor posted a blog entry Circadian Regulation,NMN, Preventing Diabetes, and Longevity - an introduction to the various clock processes affecting our cells and systems and to the relationship between epigenetics and circadian regulation. Among matters laid out there was Victor’s argument “The suggestion is that direct supplementation of NMN, which is the rate-limiting step in NAD production, could have therapeutic potential, while effectively bypassing the need for NAMPT and its potentially harmful, proinflammatory effects. ” That observation was published in January 2012, almost two years before the current focus on NMN supplementation triggered by David Sinclair’s important December 2013 paper.
This current blog entry highlights selected additional important insights and recent discoveries. Because the area of circadian rhythm-related research is so vast, I am forced to be very selective in deciding on the research to be covered here. I have possibly left out some important findings. One area I do touch seriously on is gene expression. It appears that expression of at least 10% of our genes is regulated by our biological clocks. So, the biological impacts of many things we do – including exercise and other interventions affecting aging – are highly dependent on the states of our clocks when we do them. And those actions may in turn affect the states and functioning of our clocks. Recent discovery after discovery brings home the importance of clock functions to our health and the necessity of respecting them. We are making discoveries about our mechanisms for modifying and resetting our clocks and gaining insights as to how clocks respond to metabolic conditions in cells. And a vast area of new research is just opening up related to how expression of various RNAs affect our circadian rhythms and vice-a-versa.
From the time when our original predecessors crawled out of the ocean onto land, it became important for organisms to coordinate and optimize their metabolic and other internal activities with daily rhythms of night and dark. This applied to both plants and animals. The eventual evolutionary result was the emergence of numerous biological clocks linked to master clocks in higher organisms like ourselves. Mechanisms developed for resetting, synchronizing and controlling these clocks. Reading on here, it should become clear that conforming our activities to respecting those clocks is important for maintaining our health and modulating our aging. However, our modern lifestyles involve many factors that can disrupt our regular circadian rhythms, including but not confined to: air travel over time zones, night work, irregular sleeping patterns, irregular and late-night meals, alcohol, binge eating and drinking, stimulants like caffeine, stresses, bright lights during normal dark night times, and exposure to bright TV and computer screens before bed. Even the color temperatures of our bedroom and bathroom lights may be relevant. And aging itself may be accompanied by or accelerated by clock dysregulation. That is the relevance of this blog topic.
Some basics in review
- Living biological organisms respond to multiple temporal rhythms, the science involved being Chronobiology. Chronobiological rhythms include daily, weekly, seasonal, and annual rhythms and special ones such as connected with tides and reproduction. The rhythms are coordinated by internal clocks on the cellular level, and it may be that the periodicity of one such clock in higher organisms is the entire lifespan.
- The most studied rhythms and the ones I focus on here are the daily (circadian) ones.
- “A circadian clock, or circadian oscillator, is a biochemical mechanism that oscillates with a period of 24 hours and is coordinated with the day-night cycle. Circadian clocks are the central mechanisms which drive circadian rhythms. Viewed classically, they consist of three major components: a) A central oscillator with a period of about 24 hours that keeps time, b). A series of input pathways to this central oscillator to allow entrainment of the clock, and c) A series of output pathways tied to distinct phases of the oscillator that regulate overt rhythms in biochemistry, physiology, and behavior throughout an organism(ref)”
- The clock is reset or regulated (entrained) “as the environment changes through an organism’s ability to sense external time cues of which the primary one is light. Circadian oscillators are ubiquitous in tissues of the body where they are synchronized by both endogenous and external signals to regulate transcriptional activity throughout the day in a tissue-specific manner.[1] The circadian clock is intertwined with most cellular metabolic processes and it is affected by organism aging.[2]
- the body has a number of circadian clocks and a master clock which to some extent are coordinated. “Although circadian rhythms are endogenous (“built-in”, self-sustained), they are adjusted (entrained) to the local environment by external cues called zeitgebers, commonly the most important of which is daylight(ref)”.
- “In mammals, the suprachiasmatic nucleus (SCN) of the hypothalamus constitutes the central circadian pacemaker. The SCN receives light signals from the retina and controls peripheral circadian clocks (located in the cortex, the pineal gland, the liver, the kidney, the heart, etc.). This hierarchical organization of the circadian system ensures the proper timing of physiological processes. In each SCN neuron, interconnected transcriptional and translational feedback loops enable the circadian expression of the clock genes(ref)”
- “The peripheral clocks are entrained by metabolic factors, like feeding and physical activity, while the central clock in the suprachiasmatic nucleus (SCN) of the hypothalamus is entrained by ambient conditions, such as light and to a lesser extent temperature(ref).”
As we see below, recent research challenges the above classical notions in certain respects. For example, the clocks do not necessarily respond to each other in a hierarchical manner. There appears to be clocks in every cell and the various clocks may or may not be synchronized to master clocks or to each other.
Image source “Biological clock affects the daily rhythm of many physiological processes. This diagram depicts the circadian patterns typical of someone who rises early in morning, eats lunch around noon, and sleeps at night (10 p.m.). Although circadian rhythms tend to be synchronized with cycles of light and dark, other factors – such as ambient temperature, meal times, stress and exercise – can influence the timing as well.”
Every human cell contains a 24-hour circadian clock which epigenetically regulates expression of thousands of genes.
The 2012 publication Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammalsreports on the molecular clock mechanisms within cells: “The mammalian circadian clock involves a transcriptional feedback loop in which CLOCK and BMAL1 activate the Period and Cryptochrome genes, which then feed back and repress their own transcription. We have interrogated the transcriptional architecture of the circadian transcriptional regulatory loop on a genome scale in mouse liver and find a stereotyped, time-dependent pattern of transcription factor binding, RNA polymerase II (RNAPII) recruitment, RNA expression, and chromatin states. We find that the circadian transcriptional cycle of the clock consists of three distinct phases: a poised state, a coordinated de novo transcriptional activation state, and a repressed state. Only 22% of messenger RNA (mRNA) cycling genes are driven by de novo transcription, suggesting that both transcriptional and posttranscriptional mechanisms underlie the mammalian circadian clock. We also find that circadian modulation of RNAPII recruitment and chromatin remodeling occurs on a genome-wide scale far greater than that seen previously by gene expression profiling.”
The article goes on to explain what goes on in the course of a day: “The circadian clock in mammals is cell autonomous and is composed of an autoregulatory transcriptional network with Unterlocked feedback loops (1, 2). At the core, the basic helix-loop-helix–PER-ARNT-SIM (PAS) transcriptional activators BMAL1,CLOCK, and NPAS2 activate the Period (Per1 and Per2) and Cryptochrome (Cry1 and Cry2) genes, whose transcripts and proteins slowly accumulate during the daytime (3–7). The PER and CRY proteins associate and translocate into the nucleus during the evening and physically interact with the LOCK/NPAS2:BMAL1 complex to repress their own transcription (5, 7, 8). As the PER and CRY proteins are progressively phosphorylated during the night, they are targeted for ubiquitination by specific E3 ligases and are eventually degraded by the proteasome (9–12). The waxing and waning of this transcriptional feedback loop takes ~24 hours to complete and represents the core mechanism of the circadian clock in mammals. Biochemical analysis on a small set of target genes has shown that CLOCK, BMAL1, and CRY1 bind in a diurnal manner to Regulatory regions; interact with p300 andCREB-binding protein (CBP); and are accompanied by rhythmic changes in histone H3 Lys4 trimethylation (H3K4me3) and H3 Lys9 acetylation (H3K9ac) (13–19) characteristic of active promoter regions (20–23). – Although the majority of the core components of the circadian gene network are likely known (1) and many thousands of transcripts have been shown to express circadian oscillations in various tissues (24), the genome-wide architecture of the transcriptional network regulated by the core circadian clock remains to be defined (25, 26).” —-
Most but not all circadian gene regulation is post-transcriptional.
“Although global circadian rhythms in steady-state mRNA levels have been known for a decade (35, 41, 42), here we find that only ~22% of cycling mRNA transcripts are driven by transcription (Fig. 2). Therefore, posttranscriptional regulatory events must contribute significantly to the generation of steady-state cycling mRNA levels (46, 47). Unexpectedly the most pervasive circadian regulation observed on a genome scale are rhythms in RNAPII recruitment and initiation, H3K4me3, H3K9ac, and H3K27ac, which occur at thousands of expressed genes whether or not RNA cycling was detectable. What accounts for these genome-wide circadian rhythms in RNAPII occupancy and histone modifications? Examination of the correlation between circadian transcription factor occupancy and gene expression shows that about 90% of genes bound by these factors are expressed whereas only 1 to 5% of unexpressed genes are similarly bound (table S10). These results demonstrate that gene expression per se, rather than rhythmicity of gene expression, is tightly correlated with circadian transcription factor binding. Rhythmic circadian transcription factor occupancy in turn could then be responsible for RNAPII recruitment and initiation on a genome-wide basis, which would then lead to the global rhythmic histone modifications seen here. Thus, circadian transcriptional regulators appear to be involved in the initial stages of RNAPII recruitment and initiation and the histone modifications associated with these events to set the stage for gene expression on a global scale, but additional control steps must then determine the ultimate transcriptional outputs from these sites(ref).”
Image source “Schematic representation of the histone H3 tail, the relevant post-translational modifications and the chromatin remodellers involved in circadian control. At least 10% of the genes in any given cell are expressed in a cyclic manner under circadian control mediated by cyclic chromatin modifications at the promoters of clock-controlled genes. Phosphorylation (P; serine 10 (S10)), acetylation (Ac; lysine 9/14 (K9/14)) and methylation (Me; K4 and K27) are associated with circadian transcription. Some chromatin modifiers may be directly or indirectly modulated by the circadian system (Table 1). Methylation at K4 is consistently associated with gene activation and might be crucial for circadian gene transcription and recruitment of circadian chromatin remodelling complexes87. Non-histone proteins can also undergo clock-dependent acetylation, as is the case for BMAL1 (Table 1). Several chromatin remodellers can be considered as metabolic sensors, as they use metabolites for their enzymatic function; one example is the NAD+-dependent deacetylase sirtuin 1 (SIRT1), which revealed a link between the circadian clock and cellular metabolism through its role in circadian control and its physical interaction with CLOCK. For enzymatic functions of these chromatin modifiers and references, see Table 1. See Refs 88,89 for further details. EZH2, enhancer of zeste homologue 2; FAD, flavin adenine dinucleotide, quinone form; FADH2, flavin adenine dinucleotide, hydroquinone form; HDAC, histone deacetylase; JARID1A, Jumonji/ARID domain-containing protein 1A; MLL1, mixed lineage leukaemia protein 1; MSK1, mitogen- and stress-activated protein kinase 1; NCoR1, nuclear receptor co-repressor 1; RSK2, ribosomal S6 kinase 2; SAH, S-adenosylhomocysteine; SAM, S-adenosylmethionine.”
Clock genes exercise epigenetic control of gene expression on clock-controlled genes via chromatin remodeling such as DNA methylation or histone tail modifications
The 2012 publication Circadian rhythms and memory formation: regulation by chromatin remodelingdescribes the basic mechanism through which clock genes exercise epigenetic control of gene expression on clock controlled genes via chromatin remodeling: “Epigenetic changes, such as DNA methylation or histone modification, can remodel the chromatin and regulate gene expression. Remodeling of chromatin provides an efficient mechanism of transducing signals, such as light or nutrient availability, to regulate gene expression. CLOCK:BMAL1 mediated activation of clock-controlled genes (CCGs) is coupled to circadian changes in histone modification at their promoters. Several chromatin modifiers, such as the deacetylases SIRT1 and HDAC3 or methyltransferase MLL1, have been shown to be recruited to the promoters of the CCGs in a circadian manner. Interestingly, the central element of the core clock machinery, the transcription factor CLOCK, also possesses histone acetyltransferase activity. Rhythmic expression of the CCGs is abolished in the absence of these chromatin modifiers. Recent research has demonstrated that chromatin remodeling is at the cross-roads of circadian rhythms and regulation of metabolism and aging. It would be of interest to identify if similar pathways exist in the epigenetic regulation of memory formation.”
Image source “FIGURE 1. Epigenetic regulation of gene expression by circadian clock. CLOCK can acetylate histones to induce gene expression. CLOCK interacts with MLL1 (a histone methyltransferase) and SIRT1 (a deacetylase).These epigenetic regulators can modify the chromatin according the environmental stimuli, such as nutrient availability. Furthermore, REV-ERBα, a clock-controlled gene, can cause recruitment of HDAC3 and deacetylate histones. Circadian regulation of either the expression or the activity of these epigenetic regulators determines whether the gene gets turned “ON” or “OFF.”
The circadian rhythms associated with sleep are very important for the health of humans and have been studied from multiple perspectives
There is extensive body of research literature relating to circadian rhythms, sleep, sleep disorders and related pathologies(ref). There are various types of circadian rhythm sleep disorders, such as outlined in this document from the American Academy of Sleep Medicine.
Insufficient or poor sleep can have a lasting consequence on gene expression
The 2013 publication Effects of insufficient sleep on circadian rhythmicity and expression amplitude of the human blood transcriptomehas this reported significance: “Insufficient sleep and circadian rhythm disruption are associated with negative health outcomes, but the mechanisms involved remain largely unexplored. We show that one week of insufficient sleep alters gene expression in human blood cells, reduces the amplitude of circadian rhythms in gene expression, and intensifies the effects of subsequent acute total sleep loss on gene expression. The affected genes are involved in chromatin remodeling, regulation of gene expression, and immune and stress responses. The data imply molecular mechanisms mediating the effects of sleep loss on health and highlight the interrelationships between sleep homeostasis, circadian rhythmicity, and metabolism.” The abstract of the publication is “Insufficient sleep and circadian rhythm disruption are associated with negative health outcomes, including obesity, cardiovascular disease, and cognitive impairment, but the mechanisms involved remain largely unexplored. Twenty-six participants were exposed to 1 wk of insufficient sleep (sleep-restriction condition 5.70 h, SEM = 0.03 sleep per 24 h) and 1 wk of sufficient sleep (control condition 8.50 h sleep, SEM = 0.11). Immediately following each condition, 10 whole-blood RNA samples were collected from each participant, while controlling for the effects of light, activity, and food, during a period of total sleep deprivation. Transcriptome analysis revealed that 711 genes were up- or down-regulated by insufficient sleep. Insufficient sleep also reduced the number of genes with a circadian expression profile from 1,855 to 1,481, reduced the circadian amplitude of these genes, and led to an increase in the number of genes that responded to subsequent total sleep deprivation from 122 to 856. Genes affected by insufficient sleep were associated with circadian rhythms (PER1, PER2, PER3, CRY2, CLOCK, NR1D1, NR1D2, RORA, DEC1, CSNK1E), sleep homeostasis (IL6, STAT3, KCNV2, CAMK2D), oxidative stress (PRDX2, PRDX5), and metabolism (SLC2A3, SLC2A5, GHRL, ABCA1). Biological processes affected included chromatin modification, gene-expression regulation, macromolecular metabolism, and inflammatory, immune and stress responses. Thus, insufficient sleep affects the human blood transcriptome, disrupts its circadian regulation, and intensifies the effects of acute total sleep deprivation. The identified biological processes may be involved with the negative effects of sleep loss on health, and highlight the interrelatedness of sleep homeostasis, circadian rhythmicity, and metabolism.”
Circadian epigenetic clocks determine the best times for expressions of various genes and coordination of metabolic activities. Failure to respect those times, like insufficient sleep, can lead to disorders and diseases.
The 2013 publication Epigenetic regulation of the molecular clockworkreports “Circadian rhythms control a wide range of physiological events in all organisms. Typical of our modern lifestyles is the flexibility to rest, exercise, eat, or socialize at any time of the circadian day or night; yet, these allowances correlate with rising disorders of a metabolic nature, which are thought to be mediated by changes in the molecular events underlying metabolic gene expression. Because the clock confers on gene expression changes in activity that are not necessarily related to changes in DNA sequence, the study of circadian rhythms is inseparable from epigenetics. Increasingly evident is that energy balance at the systems level relies on precise and collaborative circadian timing of epigenetic events within individual cells and tissues of the body. At the center of these rhythms resides the circadian clock machinery, a remarkably well-orchestrated transcription-translation feedback system that incorporates a fluctuating landscape of mRNA expression, protein stability, chromatin state, and metabolite abundance to keep correct time. Understanding more fully the ties that exist between cellular metabolism and the circadian clock at the epigenetic level will produce not only needed insights about circadian physiology but also novel strategies for the pharmacological and nonpharmacological treatment of metabolic disorders.”
The 2013 publication The circadian clock: a framework linking metabolism, epigenetics and neuronal function reports” ““The circadian clock machinery is responsible for biological timekeeping on a systemic level. The central clock system controls peripheral clocks through a number of output cues that synchronize the system as a whole. There is growing evidence that changing cellular metabolic states have important effects on circadian rhythms and can thereby influence neuronal function and disease. Epigenetic control has also been implicated in the modulation of biological timekeeping, and cellular metabolism and epigenetic state seem to be closely linked. We discuss the idea that cellular metabolic state and epigenetic mechanisms might work through the circadian clock to regulate neuronal function and influence disease states.”
Dysregulation of our epigenetic clocks or lifestyle behaviors that do not respect them can lead to a wide variety of disease susceptibilities and processes
Exposure to blue or ultraviolet-intensive light at the wrong time can also disrupt light-entrained rhythms and lead to pathologies, as pointed out in the 2012 blog entry Blue light, sleep, mental alertness and health.
It appears that our epigenetic clocks track if not to some extent regulate organismal aging.
There is a growing consensus that our circadian clocks have a lot to do with aging. A Pubmed search on “circadian rhythms aging” produces 2786 research publication citations. (9 more today than the same search produced yesterday). The September 2013 publication How does the body know how old it is? Introducing the epigenetic clock hypothesisreports “Animals and plants have biological clocks that help to regulate circadian cycles, seasonal rhythms, growth, development, and sexual maturity. It is reasonable to suspect that the timing of senescence is also influenced by one or more biological clocks. Evolutionary reasoning first articulated by G. Williams suggests that multiple, redundant clocks might influence organismal aging. Some aging clocks that have been proposed include the suprachiasmatic nucleus, the hypothalamus, involution of the thymus, and cellular senescence. Cellular senescence, mediated by telomere attrition, is in a class by itself, having recently been validated as a primary regulator of aging. Gene expression is known to change in characteristic ways with age, and in particular DNA methylation changes in age-related ways. Herein, I propose a new candidate for an aging clock, based on epigenetics and the state of chromosome methylation, particularly in stem cells. If validated, this mechanism would present a challenging target for medical intervention.”
Aging can lead to dysregulation of biological clock systems resulting in immune system impairment and increased disease susceptibilities.
The 2014 publication Chronobiology of the Neuroimmunoendocrine System and Agingreports: “The health maintenance depends on the preservation of the homeostatic systems, such as nervous, endocrine and immune system, and a proper communication between them. In this regard, the circadiansystem, which promotes a better functionality of physiological systems and thus well being, could be considered part of that homeostatic complex, since the neuroimmunoendocrine system possesses circadian patterns in most variables, as well as circannual or seasonal variations. With aging, an impairment of the homeostatic systems occurs and an alteration of circadian system regulation has been demonstrated. In the immune system, several function parameters, which are good markers of health and of the rate of aging, change not only with age (immunosenescence) but also throughout the day and year. Indeed, with advancing age there is a modification of immune cell circadian functionality especially in lymphocytes. Moreover, immune functions at early afternoon correspond to more aged values than at morning, especially in mature subjects (60-79 years of age). In addition, these mature men and women showed a significant impaired immune cell function, which is especially remarkable in the winter. It is noteworthy the role of immunomodulatory hormones, such as melatonin, in the regulation of biologicalrhythms and their involvement in the aging process. Furthermore, the evidence of a neuroimmune regulation of the circadian system and its disturbance with aging, highlights the importance of proinflammatory cytokines in this complex cross-talk. The biological rhythms disruption with age and some diseases (jet lag, cancer and seasonal affective disorder), could contribute increasing the immune system impairment and consequently the loss of health.”
The 2013 publication Noninvasive fractal biomarker of clock neurotransmitter disturbance in humans with dementiareports: “Human motor activity has a robust, intrinsic fractal structure with similar patterns from minutes to hours. The fractal activity patterns appear to be physiologically important because the patterns persist under different environmental conditions but are significantly altered/reduced with aging and Alzheimer’sdisease(AD). Here, we report that dementia patients, known to have disrupted circadian rhythmicity, also have disrupted fractal activity patterns and that the disruption is more pronounced in patients with more amyloid plaques (a marker of AD severity). Moreover, the degree of fractal activity disruption is strongly associated with vasopressinergic and neurotensinergic neurons (two major circadian neurotransmitters) in postmortem suprachiasmatic nucleus (SCN), and can better predict changes of the two neurotransmitters than traditional circadian measures. These findings suggest that the SCN impacts human activity regulation at multiple time scales and that disrupted fractal activity may serve as a non-invasive biomarker of SCN neurodegeneration in dementia.”
The Complex (Epigenetic) Explanation for Sleep
The “Circadian Cistrome” (aka Epigenetic explanations for the necessity of sleep): Gene expression is regulated by epigenetic mechanisms by a molecular 24-hour clock
The epigenetic explanation for why sleep is so important is probably a much more comprehensive explanation than others, but is not easy to understand. It has to do with the epigenetic regulation of genes based on circadian rhythms. The details of this have recently been worked out by Dr. Joesph Takahashi, a researcher at UT Southwestern. Takahashi and his colleagues generated the above-mentioned paper Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Sleep is important because it is important for healthy gene expression, and sustained lack of sleep can have long-lasting negative consequences relating to gene expression,
The chronobiological responses of men and women are somewhat different.
The 2014 publication Chronotype ontogeny related to genderreports: “Chronotype is an established concept designed to identify distinct phase relationships between the expression of circadian rhythms and external synchronizers in humans. Although it has been widely accepted that chronotype is subjected to ontogenetic modulation, there is no consensus on the interaction between age and gender. This study aimed to determine the relationship between age- and gender-related changes in the morningness-eveningness character in a large sample of people. A total of 14,650 volunteers were asked to complete the Brazilian version of the Horne and Östberg chronotype questionnaire. The data demonstrated that, on average, women were more morning-oriented than men until the age of 30 and there were no significant differences between men and women from 30 to 45 years of age. In contrast to the situation observed until the age of 30, women older than 45 years were more evening-oriented than men. These results suggest that the ontogenetic development of thecircadian timekeeping system is more plastic in men, as represented by the larger amplitude of chronotype changes throughout their aging process. The phase delay of adolescence and phase advance of the elderly seem to be phenomena that are more markedly present in men than in women. Thus, our data, for the first time, provide support that sharply opposes the view that there is a single path toward morningness as a function of age, regardless of gender.”
Sleep deprivation is one well-studied intervention that prevents circadian entrainment and that can lead to multiple pathologies.
In mouse pancreatic cells at least, sleep deprivation stress is associated with improper protein folding and induction of the unfolded protein response (UPR) in the endoplasmic reticulum and upregulation of apoptosis. One consequence in older animals can be hyperglycemia.
The November 2013 publication Aging and sleep deprivation induce the unfolded protein response in the pancreas: implications for metabolismreports: “Sleep disruption has detrimental effects on glucose metabolism through pathways that remain poorly defined. Although numerous studies have examined the consequences of sleep deprivation (SD) in the brain, few have directly tested its effects on peripheral organs. We examined several tissues in mice for induction of the unfolded protein response (UPR) following acute SD. In young animals, we found a robust induction of BiP in the pancreas, indicating an active UPR. At baseline, pancreata from aged animals exhibited a marked increase in a pro-apoptotic transcription factor, CHOP, that was amplified by SD, whereas BiP induction was not observed, suggesting a maladaptive response to cellular stress with age. Acute SD increased plasma glucose levels in both young and old animals. However, this change was not overtly related to stress in the pancreatic beta cells, as plasma insulin levels were not lower following acute SD. Accordingly, animals subjected to acute SD remained tolerant to a glucose challenge. In a chronic SD experiment, young mice were found to be sensitized to insulin and have improved glycemic control, whereas aged animals became hyperglycemic and failed to maintain appropriate plasma insulin concentrations. Our results show that both age and SD cooperate to induce the UPR in pancreatic tissue. While changes in insulin secretion are unlikely to play a major role in the acute effects of SD, CHOP induction in pancreatic tissues suggests that chronic SD may contribute to the loss or dysfunction of endocrine cells and that these effects may be exacerbated by normal aging.”
Five years ago, researchers at the Perelman School of Medicine, University of Pennsylvania, showed that the UPR is an adaptive response to stress induced by sleep deprivation and is impaired in the brains of old mice. Those findings suggested that inadequate sleep in the elderly, who normally experience sleep disturbances, could exacerbate an already-impaired protective response to protein misfolding that happens in aging cells. Protein misfolding and clumping is associated with many diseases such as Alzheimer’s and Parkinson’s, noted Nirinjini Naidoo, Ph.D., research associate professor in the Division of Sleep Medicine in that study.
Naidoo is also senior author of a follow-up study in Aging Cell this month that shows, for the first time, an effect of sleep deprivation on the UPR in peripheral tissue, in this case, the pancreas. They showed that stress in pancreatic cells due to sleep deprivation may contribute to the loss or dysfunction of these cells important to maintaining proper blood sugar levels, and that these functions may be exacerbated by normal aging. — “The combined effect of aging and sleep deprivation resulted in a loss of control of blood sugar reminiscent of pre-diabetes in mice,” says Naidoo. “We hypothesize that older humans might be especially susceptible to the effects of sleep deprivation on the disruption of glucose homeostasis via cell stress.” — Working with Penn colleague Joe Baur, Ph.D., assistant professor of Physiology, Naidoo started a collaboration to look at the relationship of sleep deprivation, the UPR, and metabolic response with age. Other researchers had suggested that the death of beta cells associated with type 2 diabetes may be due to stress in a cell compartment called the endoplasmic reticulum (ER). The UPR is one part of the quality control system in the ER, where some proteins are made. Knowing this, Naidoo and Baur asked if sleep deprivation (SD) causes ER stress in the pancreas, via an increase in protein misfolding, and in turn, how this relates to aging. The team examined tissues in mice for cellular stress following acute SD, and they also looked for cellular stress in aging mice. Their results show that both age and SD combine to induce cellular stress in the pancreas.
Older mice fared markedly worse when subjected to sleep deprivation. Pancreas tissue from older mice or from young animals subjected to sleep deprivation exhibited signs of protein misfolding, yet both were able to maintain insulin secretion and control blood sugar levels. Pancreas tissue from acutely sleep-deprived aged animals exhibited a marked increase in CHOP, a protein associated with cell death, suggesting a maladaptive response to cellular stress with age that was amplified by sleep deprivation. Acute sleep deprivation caused increased plasma glucose levels in both young and old animals. However, this change was not overtly related to stress in beta cells, since plasma insulin levels were not lower following acute lack of sleep. — Accordingly, young animals subjected to acute sleep deprivation remained tolerant to a glucose challenge. In a chronic sleep deprivation experiment, young mice were sensitized to insulin and had improved control of their blood sugar, whereas aged animals became hyperglycemic and failed to maintain appropriate plasma insulin concentrations.
While changes in insulin secretion are unlikely to play a major role in the acute effects of SD, cellular stress in pancreatic tissue suggests that chronic SD may contribute to the loss or dysfunction of endocrine cells, and that these effects may be exacerbated by normal aging, say the researchers.”
Newer techniques of genetic and epigenetic profile and computer mathematical systems modeling are being applied to study the impacts of circadian rhythms on gene expression
The 2013 publication Circadian rhythms investigated on the cellular and molecular levelsreports “Investigations on circadian rhythms have markedly advanced our understanding of health and disease with the advent of high-throughput technologies like microarrays and epigenetic profiling. They elucidated the multi-level behaviour of interactive oscillations from molecules to neuronal networks and eventually to processes of learning and memory in an impressive manner. The small-world topology of synchronized firing through neuron-neuron and neuron-glia gap junctions is discussed as a mathematical approach to these intensively studied issues. It has become evident that, apart from some disorders caused by gene mutations, the majority of disorders originating from disturbances of rhythms arise from environmental influences and epigenetic changes. In this context, it was mandatory to think of and devise experiments on temporary scales, which exponentially increased the volumes of data obtained from time-series and rapidly became prohibitive of manual inspection. Therefore, more and more sophisticated mathematical algorithms have been developed to identify rhythmic expression of genes and to find coexpression by their clustering. It is expected that disturbed rhythmic behaviour in mental disorders is reflected in altered oscillatory behaviour of gene expression.”
The networks that relate clock gene, circadian rhythms and gene activation in organs and individual networks and within neurons themselves are very complex and are being actively studied using computer and mathematical modeling approaches.
The interactions among clocks is complex and not hierarchical
It has been thought that body clocks function in a somewhat hierarchical manner with one or a few “master” clocks controlling the other ones. That model is probably not correct with different clocks being able to come to the fore and control others depending on state, state dynamics, and circumstances. At least this appears to be the case in the case of clock neurons in the fruit fly. The 2014 publication The DrosophilaCircadian Clock Is a Variably Coupled Network of Multiple Peptidergic Unitsreports “Daily rhythms in behavior emerge from networks of neurons that express molecular clocks. Drosophila’s clock neuron network consists of a diversity of cell types, yet is modeled as two hierarchically organized groups, one of which serves as a master pacemaker. Here, we establish that the fly’s clock neuron network consists of multiple units of independent neuronal oscillators, each unified by its neuropeptide transmitter and mode of coupling to other units. Our work reveals that the circadian clock neuron network is not orchestrated by a small group of master pacemakers but rather consists of multiple independent oscillators, each of which drives rhythms in activity.” The Editor’s Summary states “Circadian rhythms in the fruit fly Drosophilia are driven by neurons in the brain. Yao and Shafer (p. 1516) analyzed different sets of neurons that can drive circadian rhythms. Manipulating the period of each set of neurons separately revealed that when the various clock signals were fairly consistent, the fly showed a robust circadian rhythm. But when the various clock signals were seriously out of sync with one another, the fly was oblivious to the day-night cycle.”
The same research as reported in a University of Michigan 2014 press release is commented on as follows: “— Circadian clock like an orchestra with many conductors — Now, new findings from Shafer and U-M doctoral student Zepeng Yao challenge the prevailing wisdom about how our body clocks are organized, and suggest that interactions among neurons that govern circadian rhythms are more complex than originally thought. — Yao and Shafer looked at the circadian clock neuron network in fruit flies, which is functionally similar to that of mammals, but at only 150 clock neurons is much simpler. Previously, scientists thought that a master group of eight clock neurons acted as pacemaker for the remaining 142 clock neurons — think of a conductor leading an orchestra — thus imposing the rhythm for the fruit fly circadian clock. It is thought that the same principle applies to mammals. — Interactions among clock neurons determine the strength and speed of circadian rhythms, Yao says. So, when researchers genetically changed the clock speeds of only the group of eight master pacemakers they could examine how well the conductor alone governed the orchestra. They found that without the environmental cues, the orchestra didn’t follow the conductor as closely as previously thought. — Some of the fruit flies completely lost sense of time, and others simultaneously demonstrated two different sleep cycles, one following the group of eight neurons and the other following some other set of neurons. “The finding shows that instead of the entire orchestra following a single conductor, part of the orchestra is following a different conductor or not listening at all,” Shafer said. The findings suggest that instead of a group of master pacemaker neurons, the clock network consists of many independent clocks, each of which drives rhythms in activity. Shafer and Yao suspect that a similar organization will be found in mammals, as well. — “
Our old friend/enemy IGF-1 is implicated in sleep disruption, at least in the fruit fly. Further, inhibition of IGF-1 signaling and TOR may be an intervention that enhances sleep
Followers of the aging sciences know that IGF-1 signaling is essential in early animal development and yet plays a strong pro-aging and pro-disease role later in life. Indeed in many animals, inhibition of IGF-1 signaling in mature animals via TOR inhibition is a proven strategy for modest life extension That is how rapamycin extends life. Here is another angle: inhibition of IGF-1 signaling can lead to better sleep – at least in the fruit fly.
The 2014 publication Lowered Insulin Signalling Ameliorates Age-Related Sleep Fragmentation in Drosophila reports “Sleep fragmentation, particularly reduced and interrupted night sleep, impairs the quality of life of older people. Strikingly similar declines in sleep quality are seen during ageing in laboratory animals, including the fruit fly Drosophila. We investigated whether reduced activity of the nutrient- and stress-sensing insulin/insulin-like growth factor (IIS)/TOR signalling network, which ameliorates ageing in diverse organisms, could rescue the sleep fragmentation of ageing Drosophila. Lowered IIS/TOR network activity improved sleep quality, with increased night sleep and day activity and reduced sleep fragmentation. Reduced TOR activity, even when started for the first time late in life, improved sleep quality. The effects of reduced IIS/TOR network activity on day and night phenotypes were mediated through distinct mechanisms: Day activity was induced by adipokinetic hormone, dFOXO, and enhanced octopaminergic signalling. In contrast, night sleep duration and consolidation were dependent on reduced S6K and dopaminergic signalling. Our findings highlight the importance of different IIS/TOR components as potential therapeutic targets for pharmacological treatment of age-related sleep fragmentation in humans.” Further, “Sleep is essential for human health, but the quality of this fundamental physiological process declines with age and reduces quality of life. We therefore investigated the mechanisms by which ageing impairs sleep. We used the fruit fly Drosophila, whose sleep has many features in common with that of humans, including the age-related decline in quality. We examined the role of the insulin/IGF (IIS) and TOR signaling network, which has an evolutionarily conserved role in ageing. We found that flies with reduced IIS activity had improved sleep quality at night and higher activity levels by day. Importantly, day activity and night sleep were regulated through distinct mechanisms—day activity by the key IIS transcription factor dFOXO, adipokinetic hormone, and octopaminergic signalling—whereas night sleep was mediated through TOR and dopaminergic signalling. Surprisingly, acute inhibition of TOR, by rapamycin, even in old flies, improved sleep quality, suggesting that age-related sleep decline is reversible even after it has commenced. Given the high evolutionarily “conservation of IIS and TOR function, our results implicate potential therapeutic targets to improve sleep quality in humans.”
Natural expression of the melatonin hormone responds to the circadian cycle and exogenous hormone supplementation can be used to regulate the sleep cycle.
Melatonin is mentioned here because it has been much studied in relationship to circadian sleep rhythms (ref) and is widely used as a supplement. A Pubmed search on “melatonin circadian rhythm” today produces 5765 research article citations, more tomorrow. An entire blog entry could be devoted to its actions, and perhaps should be.
For the moment I mention only that here are many interesting sides to the melatonin story, for example, how circadian regulation or external taking of melatonin might affect dental hygiene and control dental biofilm and plaque formation and the formation of cavities. Relative to this, there is a 2014 review article: Melatonin in the oral cavity: physiological and pathological implications: “BACKGROUND AND OBJECTIVES: The purpose of this article was to summarize what is known about the function of melatonin in the oral cavity. MATERIAL AND METHODS: Databases were searched for the relevant published literature to 30 November, 2013. The following search items were used in various combinations: melatonin, gingiva, periodontium, inflammation, herpes, alveolar bone, periodontal ligament, dental implants, xerostomia, methacrylate, chlorhexidine, cancer. The literature uncovered is summarized herein. RESULTS: Salivary melatonin levels exhibit a circadian rhythm with highest values at night. Melatonin has both receptor-mediated and receptor-independent actions in cells of the oral cavity. Melatonin is released into the saliva by the acinar cells of the major salivary glands and via the gingival fluid. Functions of melatonin in the oral cavity are likely to relate primarily to its anti-inflammatory and antioxidant activities. These actions may suppress inflammation of the gingiva and periodontium, reduce alveolar bone loss, abrogate herpes lesions, enhance osteointegration of dental implants, limit oral cancer, and suppress disorders that have a free radical component. Sublingual melatonin tablets or oralmelatonin sprays and topica melatonin-containing gel, if used on a regular basis, may improve overall oral health and reduce mucosal lesions. CONCLUSION: Collectively, the results indicate that endogenously-produced and exogenously-appliedmelatonin are beneficial to the oral cavity.’
Another interesting side of the melatonin-circadian story relates to early-stage human development. The 2014 review article Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology reports: “BACKGROUND: Research within the last decade has shown melatonin to have previously-unsuspected beneficial actions on the peripheral reproductive organs. Likewise, numerous investigations have documented that stable circadian rhythms are also helpful in maintaining reproductive health. The relationship of melatonin and circadian rhythmicity to maternal and fetal health is summarized in this review. METHODS: Databases were searched for the related published English literature up to 15 May 2013. The search terms used in various combinations included melatonin, circadian rhythms, biological clock, suprachiasmatic nucleus, ovary, pregnancy, uterus, placenta, fetus, pre-eclampsia, intrauterine growth restriction, ischemia-reperfusion, chronodisruption, antioxidants, oxidative stress and free radicals. The results of the studies uncovered are summarized herein. RESULTS: Both melatonin and circadian rhythms impact reproduction, especially during pregnancy. Melatonin is a multifaceted molecule with direct free radical scavenging and indirect antioxidant activities. Melatonin is produced in both the ovary and in the placenta where it protects against molecular mutilation and cellular dysfunction arising from oxidative/nitrosative stress. The placenta, in particular, is often a site of excessive free radical generation due to less than optimal adhesion to the uterine wall, which leads to either persistent hypoxia or intermittent hypoxia and reoxygenation, processes that cause massive free radical generation and organ dysfunction. This may contribute to pre-eclampsia and other disorders which often complicate pregnancy. Melatonin has ameliorated free radical damage to the placenta and to the fetus in experiments using non-human mammals. Likewise, the maintenance of a regular maternal light/dark and sleep/wake cycle is important to stabilize circadianrhythms generated by the maternal central circadian pacemaker, the suprachiasmatic nuclei. Optimalcircadian rhythmicity in the mother is important since her circadian clock, either directly or indirectly via the melatonin rhythm, programs the developing master oscillator of the fetus. Experimental studies have shown that disturbed maternal circadian rhythms, referred to as chronodisruption, and perturbedmelatonin cycles have negative consequences for the maturing fetal oscillators, which may lead to psychological and behavioral problems in the newborn. To optimize regular circadian rhythms and prevent disturbances of the melatonin cycle during pregnancy, shift work and bright light exposure at night should be avoided, especially during the last trimester of pregnancy. Finally, melatonin synergizes with oxytocin to promote delivery of the fetus. Since blood melatonin levels are normally highest during the dark period, the propensity of childbirth to occur at night may relate to the high levels of melatonin at this time which work in concert with oxytocin to enhance the strength of uterine contractions. CONCLUSIONS: A number of conclusions naturally evolve from the data summarized in this review: (i)melatonin, of both pineal and placental origin, has essential functions in fetal maturation and placenta/uterine homeostasis; (ii) circadian clock genes, which are components of all cells including those in the peripheral reproductive organs, have important roles in reproductive and organismal (fetal and maternal) physiology; (iii) due to the potent antioxidant actions of melatonin, coupled with its virtual absence of toxicity, this indoleamine may have utility in the treatment of pre-eclampsia, intrauterine growth restriction, placental and fetal ischemia/reperfusion, etc. (iv) the propensity for parturition to occur at night may relate to the synergism between the nocturnal increase in melatonin and oxytocin.”
New molecular approaches are being discovered for resetting and regulating circadian clock functions.
For example, it appears that that the enzyme casein kinase 1epsilon (CK1epsilon) controls ease of entrainment, that is, how easily the body’s clockwork can be adjusted or reset by environmental cues such as light and temperature. The 2014 publication A Novel Mechanism Controlling Resetting Speed of the Circadian Clock to Environmental Stimulirelates to clock phase resetting and clock speeds. It reports: “Many aspects of mammalian physiology are driven through the coordinated action of internal circadian clocks. Clock speed (period) and phase (temporal alignment) are fundamental to an organism’s ability to synchronize with its environment. In humans, lifestyles that disturb these clocks, such as shift work, increase the incidence of diseases such as cancer and diabetes. Casein kinases 1δ and ε are closely related clock components implicated in period determination. However, CK1δ is so dominant in this regard that it remains unclear what function CK1ε normally serves. Here, we reveal that CK1ε dictates how rapidly the clock is reset by environmental stimuli. Genetic disruption of CK1ε in mice enhances phase resetting of behavioral rhythms to acute light pulses and shifts in light cycle. This impact of CK1ε targeting is recapitulated in isolated brain suprachiasmatic nucleus and peripheral (lung) clocks during NMDA- or temperature-induced phase shift in association with altered PERIOD (PER) protein dynamics. Importantly, accelerated re-entrainment of the circadian system in vivo and in vitro can be achieved in wild-type animals through pharmacological inhibition of CK1ε. These studies therefore reveal a role for CK1ε in stabilizing the circadian clock against phase shift and highlight it as a novel target for minimizing physiological disturbance in shift workers.”
A relatively new and only partially explored frontier of circadian-related research involves the roles of various species of RNAs – coding RNAs, micro RNAs, long non-coding RNAs, alternative-splice RNAs, lincRNAs, circular RNAs, etc. It appears both that there is both circadian control of expression of certain RNAs, and that certain RNAs can affect clock gene expression and circadian rhythms.
Initial research based on the fruit fly model suggests that there is significant circadian clock control of expression on non-coding RNAs, as well as control of alternative splicing and RNA editing.
The 2012 e- publication Deep sequencing the circadian and diurnal transcriptome of Drosophila brain reports: “Eukaryotic circadian clocks include transcriptional/translational feedback loops that drive 24-h rhythms of transcription. These transcriptional rhythms underlie oscillations of protein abundance, thereby mediating circadian rhythms of behavior, physiology, and metabolism. Numerous studies over the last decade have used microarrays to profile circadian transcriptional rhythms in various organisms and tissues. Here we use RNA sequencing (RNA-seq) to profile the circadian transcriptome of Drosophila melanogaster brain from wild-type and period-null clock-defective animals. We identify several hundred transcripts whose abundance oscillates with 24-h periods in either constant darkness or 12 h light/dark diurnal cycles, including several noncoding RNAs (ncRNAs) that were not identified in previous microarray studies. Of particular interest are U snoRNA host genes (Uhgs), a family of diurnal cycling noncoding RNAs that encode the precursors of more than 50 box-C/D small nucleolar RNAs, key regulators of ribosomal biogenesis. Transcriptional profiling at the level of individual exons reveals alternative splice isoforms for many genes whose relative abundances are regulated by either period or circadian time, although the effect of circadian time is muted in comparison to that of period. Interestingly, period loss of function significantly alters the frequency of RNA editing at several editing sites, suggesting an unexpected link between a key circadian gene and RNA editing. We also identify tens of thousands of novel splicing events beyond those previously annotated by the modENCODE Consortium, including several that affect key circadian genes. These studies demonstrate extensive circadian control of ncRNA expression, reveal the extent of clock control of alternative splicing and RNA editing, and provide a novel, genome-wide map of splicing in Drosophila brain.”
Micro RNAs play important roles in regulating circadian functions
The 2013 publication Micro-managing the circadian clock: The role of microRNAs in biological timekeepingreports: “Evolved under the selective pressures of a 24-h world, circadian timekeeping mechanisms are present in virtually all living organisms to coordinate daily rhythms in physiology and behavior. Until recently, the circadian clock was modeled as simple, interlocked transcription-translation feedback loops driving rhythms in gene expression of a handful of core clock genes. However, it has become evident that circadian clock regulation is immensely more complex than once thought and involves posttranscriptional, translational and posttranslational mechanisms. In particular, there has been a growing awareness of the vital role played by microRNAs (miRNAs) in regulating various aspects of circadian clock function. In this review, we will summarize our current knowledge of miRNA-dependent regulation of the circadian timing system in multiple organisms, including flies, mammals and higher plants. We will also discuss future perspectives for research on the role of miRNAs and noncoding RNAs in circadian regulation of health and disease.”
Another popular model besides the fruit fly for studying circadian rhythms with respect to gene expression is the zebra fish.
The 2014 publication The light-induced transcriptome of the zebrafish pineal gland reveals complex regulation of the circadian clockwork by lightreports: “Light constitutes a primary signal whereby endogenous circadian clocks are synchronized (‘entrained’) with the day/night cycle. The molecular mechanisms underlying this vital process are known to require gene activation, yet are incompletely understood. Here, the light-induced transcriptome in the zebrafish central clock organ, the pineal gland, was characterized by messenger RNA (mRNA) sequencing (mRNA-seq) and microarray analyses, resulting in the identification of multiple light-induced mRNAs. Interestingly, a considerable portion of the molecular clock (14 genes) is light-induced in the pineal gland. Four of these genes, encoding the transcription factors dec1, reverbb1, e4bp4-5 and e4bp4-6, differentially affected clock- and light-regulated promoter activation, suggesting that light-input is conveyed to the core clock machinery via diverse mechanisms. Moreover, we show that dec1, as well as the core clock gene per2, is essential for light-entrainment of rhythmic locomotor activity in zebrafish larvae. Additionally, we used microRNA (miRNA) sequencing (miR-seq) and identified pineal-enhanced and light-induced miRNAs. One such miRNA, miR-183, is shown to downregulate e4bp4-6 mRNA through a 3′UTR target site, and importantly, to regulate the rhythmic mRNA levels of aanat2, the key enzyme in melatonin synthesis. Together, this genome-wide approach and functional characterization of light-induced factors indicate a multi-level regulation of the circadian clockwork by light.”
My comment: There seems to be agreement that the complex regulatory interactions observed in the fruit fly and the zebrafish situations are very likely to be evolutionarily conserved in mammals and are also likely to be significantly more complex in us. But so far as far as I have looked, these are yet to be thoroughly investigated and understood as far as humans are concerned. We are only at an early stage of understanding how circadian rhythms affect gene expression in humans, how RNA species affect circadian patterns. And how circadian patterns impact on expression of RNA species..
This paper published last month summarizes the situation as far as longncRNAs are concerned: Regulation of metabolism by long, non-coding RNAs. “Our understanding of genomic regulation was revolutionized by the discovery that the genome is pervasively transcribed, giving rise to thousands of mostly uncharacterized non-coding ribonucleic acids (ncRNAs). Long, ncRNAs (lncRNAs) have thus emerged as a novel class of functional RNAs that impinge on gene regulation by a broad spectrum of mechanisms such as the recruitment of epigenetic modifier proteins, control of mRNA decay and DNA sequestration of transcription factors. We review those lncRNAs that are implicated in differentiation and homeostasis of metabolic tissues and present novel concepts on how lncRNAs might act on energy and glucose homeostasis. Finally, the control ofcircadianrhythm by lncRNAs is an emerging principles of lncRNA-mediated gene regulation.”
This blog entry has been a sampler of a variety of recent research relating to circadian rhythms. Whole new blog entries could be devoted to some of the subtopics such as rhythms related to gene expression, ncRNAs, melatonin and relationships among clocks. And there is the whole field of plant circadian rhythms. I expect my colleagues and I will report further on some of these topics in the near future.
By James P Watson with editorial assistance and comments by Vince Giuliano
This is Part 3 of 3 in the series The View from the Telomere end of the Chromsome. Because of the length of the telomere story, we divided this discussion into three parts. Part 1 relates to a number of more-practical and less-technical topics: the history of telomere biology, telomere length testing, telomerase Inhibitors for cancer, and supplements that activate telomerase and their possible roles for inhibiting or preventing cancers. Part 2 deals in finer detail with newer discoveries related to the molecular biology of telomeres, Sections 1 – 9. This part 3 covers Sections 10-17 and a concluding discussion related to some of the implications for a Grand Unified Theory of aging and biology, as viewed from the ends of a chromosome.
The 1994 telomere-telomerase story, and the 2014 one
Introductory comment by Vince: This particular blog series is for those of you seriously interested in the science related to telomeres, above and beyond the telomere romance of two decades ago. This involves tough reading and some times even tougher understanding. I clearly recall the days in 1994 when I could relate telomere-telomerase story over a glass of wine to an artist or massage-therapist friend and have the friend get it: “Telomeres are like inert shoelace tips at the end of chromosomes that grow a little shorter every time a cell divides. They get shorter and shorter until they can’t do their job and drive a cell into senescence or death. They are like clocks counting down. Telomerase is an enzyme that pastes telomeres back on again. Doing this can run the aging clocks backwards and offers a hope for extending life.” The 2014 story told in these blog entries is vastly more complicated and makes that 1994 version seem incredibly naïve as well as downright wrong. I can’t possibly explain the story told here to most of my MD friends even over a whole case of wine, let alone to my artist friends. Yet the 2014 story is incredibly rich and illustrative of the multiple checks and balances and feedback loops in human biology. And it illustrates that there is no center to biology. You can start anywhere and get to most anywhere else. So, for those of you with the patience, we continue to tell that story here. We pick up where Part 2 left off with item 10. And the last part related to a GUT of biology is definitely interesting.
10. Regulatory Elements Prevent Silencing of Subtelomeric Genes
Using Transgene Experiments for Basic Science Discoveries
Since subtelomeric genes are vulnerable to the Telomeric Position Effect (TPE) which results in unwanted gene silencing, several complex epigenetic mechanisms have evolved to counteract this. The next four Sections here specifically review these, but as an introduction, I must explain that most of these discoveries were made using transgene experiments of genes spliced into telomeric and subtelomeric DNA to study how DNA is silenced in this area of the chromosome. As you may know, the word “transgene” refers to the procedure that most of us refer to as “genetic engineering” or recombinant DNA technology. With genetic engineering, a piece of non-native DNA (the transgene) is introduced into the genome. The location that the transgene is spliced into the DNA is random and not controllable. When the transgene is spliced into the telomeric or subtelomeric DNA, the expression of the transgene is very low or inconsistent, due to epigenetic silencing of the transgene. Once researchers discovered this, they used this as a laboratory tool to figure out why this occurred. Thus came the discoveries detailed below.
Epigenetic regulators
Now that we are about to explore even more “epigenetic tricks” up Mother Nature’s sleeve to counter-act the Telomere Position Effect (TPE), you might want to refer back to Sections 4, 5 and 6 in Part 2 of this series for a more detailed explanation of the TPE. Using transgene experiments, the following methods for counteracting the TPE were discovered. This list of new epigenetic “tricks” are called Epigenetic Regulators. They include the following:
1. MARs – Matrix attachment regions
2. UCOE – Ubiquitous chromatin opening element
3. Insulators – Example: the chicken beta-globin 5′ hypersensitive site 4 (cHS4)
4. STARs – Stabilizing Anti-repressor elements – I am not going to go over this, though it is yet another one.
The next few sections will explain, MARs, UCOEs and Insulators
The 2013 publication Epigenetic regulatory elements associate with specific histone modifications to prevent silencing of telomeric genesreports: “In eukaryotic cells, transgene expression levels may be limited by an unfavourable chromatin structure at the integration site. Epigenetic regulators are DNA sequences which may protect transgenes from such position effect. We evaluated different epigenetic regulators for their ability to protect transgene expression at telomeres, which are commonly associated to low or inconsistent expression because of their repressive chromatin environment. Although to variable extents, matrix attachment regions (MARs), ubiquitous chromatin opening element (UCOE) and the chicken cHS4 insulator acted as barrier elements, protecting a telomericdistal transgene from silencing. MARs also increased the probability of silent gene reactivation in time-course experiments. Additionally, all MARs improved the level of expression in non-silenced cells, unlike other elements. MARs were associated to histone marks usually linked to actively expressed genes, especially acetylation of histone H3 and H4, suggesting that they may prevent the spread of silencing chromatin by imposing acetylation marks on nearby nucleosomes. Alternatively, an UCOE was found to act by preventing deposition of repressive chromatin marks. We conclude that epigenetic DNA elements used to enhance and stabilize transgene expression all have specific epigenetic signature that might be at the basis of their mode of action.”
11. Insulators -The partitioners of the genome
This is something new that I had not learned about before. An “insulator” is a barrier element that partitions the genome into discreet chromatin domains. Specifically, an insulator may have an enhancer-blocking activity or a barrier activity. Here is an illustration of these two functions of insulators:
“Schematic overview of the domain model of a linear genome, highlighting insulator locations. (A) An active chromatin domain is flanked by heterochromatic regions. Insulator positions are indicated at the domain boundaries (where they can mediate border or barrier function of insulators) as well as within the active domain (where they can mediate enhancer-blocking function). It is not known whether both functions are established by similar or different mechanisms. (B) One aspect of insulator function is to organise chromatin looping by promoting contacts between insulators or with other genomic structures. Depending on the linear and three-dimensional arrangement, looping may interfere with enhancer-promoter interactions (thus mediating the enhancer-blocking function of insulators), resulting in an inactive gene (pink gene and promoter), or it may assist in increasing enhancer-promoter contacts, resulting in an active gene (green gene and promoter on right). Gene activation can also be achieved by direct enhancer-promoter interactions (green gene on left) that can occur independently of the presence of an insulator. Insulators are also found between tandem promoters positioned in a head-to-head orientation ensuring that both promoters can be regulated individually.”
In the first case in the above diagram, an insulator can interfere with the enhancer-promoter communication when interposed between them. This prevents the spread of repressive heterochromatin over adjacent euchromatin domains. Thus, insulators have been shown to confer stability to the transgene expression over time and are seen as promising tools to increase the safety of gene therapy vectors.
Numerous insulators have been identified in different species. The most well known insulator is cHS4, which is a potent insulator in chickens. It combines the enhancer-blocking and barrier activities of insulators. This is another epigenetic feature that prevents subtelomeric gene silencing due to the TPE. Here is an illustration of the cHS4 insulator:
“A model for chromatin barrier function of insulator and the histone code. The cHS4 insulator binding proteins USF1/USF2 (USF) recruit the histone acetyltransferases (HATs) PCAF and P300/CBP (H3 and H4 acetylation) and methyltransferase SET7/9 (H3 lysine 4 methylation), which form a histone hyperacetylation- and H3 lysine 4 methylation-enriched chromatin boundary to block the spread of adjacent heterochromatin structure 8, 42. The insulator perhaps recruit other putative protein factors to form a barrier complex to prevent heterochromatin spreading. Ac and Me are abbreviations of acetyl and methyl groups, respectively. Especially, Me (blue) mean methylation of H3 lysine 4, which are active chromatin marks. Other Me mean repressive chromatin marks such as methylation of H3 lysine 9.”
12. MARs – Attachement Points for DNA to the Nuclear Cytoskeleton
“Matrix attachment regions” and scaffold attachment regionsare areas of A + T rich DNA of variable length that serve as “anchoring points’ for DNA to attach to the nuclear skeleton. They are also called scaffold-attachment regions (SARs). MARs are thought to organize eukaryotic chromatin into distinct regulatory domains by the formation of 50-200 kb structural loops. Here are two illustrations of MARs.
“A proposed model for the selective use of S/MAR for transcription/replication regulation. The left panel shows a gene located on the loop with a S/MAR in close proximity. When functional demands require the specific association of this gene with the transcriptional machinery located on the nuclear matrix, the S/MAR moves the gene to the nuclear matrix, thereby initiating gene expression (center panel). Following initiation, the gene is pulled in through the transcriptional machinery, thus completing the process (right panel). There are two types of S/MARs. Functional S/MARs serve as mediators to bring genes onto the nuclear matrix. Structural S/MARs serve as anchors, which are less dynamic compared with functional S/MARs.”
“S/MAR-functions: constitutive and facultative. A chromatin domain with constitutive S/MARs at its termini (I). When functional demands require the specific translocation of the constituent gene to the matrix, facultative S/MARs responds to topological changes which are initiated by the association of transcription factors (TF) and supported by histone acetylation. Topological changes are propagated once the gene is pulled through the transcriptional machinery (II). Transcription is terminated (III) followed by dissociation of the transcription complex (IV)”
Transcription factor genes contain a significantly higher portion of MARs. As a result, when researchers find a gene with a nearby MARs, it emphasizes its functional importance. Also, when a nuclear scaffold/matrix module is included in a genetic vector such as an episome, if the episomal MAR is attached to a transcription unit, the episome will be maintained and the expression not lost over time. Thus, a MAR increases the stability of a transgene injected into a cell. It is estimated that 64,000 MARs may be present in the human genome (plus an additional 10,000 replication foci). Little sequence homology exists, however, and as a consequence, only a minor fraction have met the official criteria for a MARs. These epigenetic features are evolutionarily conserved. The chicken lysozyme MARs was one of the first seen which demonstrated increased and sustained transgene expression. In humans, the MAR 1-68 and X-29 are two of the most well understood MARs(ref). In mice, the MAR S4 is one of the most well understood matrix attachment regions. When you add a MARs to an expression vector, three things happen:
a. The number of stably expressing clones increases
b. The expression of the transgene is enhanced
c. The variability in gene expression within a polyclonal cell population is reduced
In summary, MARs appear to increase gene expression, but the exact mechanism is still not well understood. This is another epigenetic feature that helps keep subtelomeric DNA from being silenced by the TPE.
13. UCOEs - A Promoter-based method of stabilizing gene expression
Ubiquitous chromatin opening elements (UCOEs) are regulatory elements derived from promoters that contain CpG islands (remember that only about half of genes have CpG islands) They appear to be present mainly in housekeeping genes. It appears that UCOEs have chromatin remodeling function. Here is an illustration of a UCOE:
“pEPI-EGFP and derivatives. (a) Original episomal vector pEPI-EGFP harboring an active transcription unit running into the S/MAR sequence. (b) The ubiquitous chromatin-opening element (UCOE) from the HNRPA2B1-CBX3 locus was introduced upstream the CMV promoter resulting in pEPI-UCOE. (c) The chicken β-globin insulator sequence cHS4 was introduced downstream and in opposite direction to the S/MAR resulting in pEPI-HS4opp. (d) Episomal maintenance and cellular replication was ascertained in Southern blot analysis. Complete digestion with MboI indicates cellular replication (lane 1), as shown for all three pEPI derivatives. C: Plasmid DNA as control; 1: DNA digested with MboI and plasmid single cutter; 2: DNA digested with DpnI and plasmid single cutter. In no case, we detected hybridization to chromosomal DNA.”
14. HDAC5 and Telomeres – Required for maintaining normal telomeres in certain cancer cells
The publicationA new role for histone deacetylase 5 in the maintenance of long telomeresreports: “Telomeres are major regulators of genome stability and cell proliferation. A detailed understanding of the mechanisms involved in their maintenance is of foremost importance. Of those, telomere chromatin remodeling is probably the least studied; thus, we intended to explore the role of a specific histone deacetylase on telomere maintenance. We uncovered a new role for histone deacetylase 5 (HDAC5) in telomere biology. We report that HDAC5 is recruited to the long telomeres of osteosarcoma- and fibrosarcoma-derived cell lines, where it ensures proper maintenance of these repetitive regions. Indeed, depletion of HDAC5 by RNAi resulted in the shortening of longer telomeres and homogenization of telomere length in cells that use either telomerase or an alternative mechanism of telomere maintenance. Furthermore, we present evidence for the activation of telomere recombination on depletion of HDAC5 in fibrosarcoma telomerase-positive cancer cells. Of potential importance, we also found that depletion of HDAC5 sensitizes cancer cells with long telomeres to chemotherapeutic drugs. Cells with shorter telomeres were used to control the specificity of HDAC5 role in the maintenance of long telomeres. HDAC5 is essential for the length maintenance of long telomeres and its depletion is required for sensitization of cancer cells with long telomeres to chemotherapy.”
15. How Caloric Restriction and p53 can activate Telomere Maintenance Genes and DNA repair genes
Summary: Caloric restriction mimetics such as resveratrol and 2-Deoxyglucose have been shown to activate both telomere maintenance genes and DNA repair genes. These are activated via p53, using a DNA binding sequence found on the 5′ flanking regions of genes for telomere maintenance and DNA repair that do not have TATA-box or TATA-like elements. The most common binding motif is a duplicated GGAA motif or a GC box. The mechanism by which this occurs is the binding of p53 to these sites. Caloric restriction appears to activate these genes via p53 phosphorylation, which is probably induced via the inhibition of cAMP phosphodiesterase, which in turn activates AMPK pathways.
16. Relationship of telomere dysfunction and mitochondrial dysfunction
The “Axis of Aging” – Telomere Dysfunction and Mitochondrial Dysfunction are linked via DDR-activated p53 pathway
The 2012 publication Axis of ageing: telomeres, p53 and mitochondriareports: “Progressive DNA damage and mitochondrial decline are both considered to be prime instigators of natural ageing. Traditionally, these two pathways have been viewed largely in isolation. However, recent studies have revealed a molecular circuit that directly links DNA damage to compromised mitochondrial biogenesis and function via p53. This axis of ageing may account for both organ decline and disease development associated with advanced age and could illuminate a path for the development of relevant therapeutics.”
The “Axis of Aging” consists of the link between telomere shortening and mitochondrial dysfunction is now well established and explains why mitochondrial dysfunction is a universal “signature of gene expression change” with aging, as well as a universal “signature of metabolic change” with aging. Specifically, telomere dysfunction due to oxidative stress, genotoxic stress, radiation, UV, or cell divisions can result in telomere shortening and/or uncapping of Shelterin proteins, resulting in the triggering of the DNA damage response (DDR). The DDR triggers p53 activation which can then inhibit PGC-1, resulting in the inhibition of gene transcription for nuclear-encoded mitochondrial proteins. Thus, this mechanism can induce high mitochondrial ROS generation, Warburg-type metabolism, increased oxidative stress leading to further negative effects such as DNA damage. DNA damage can then lead to PARP-1 activation, which then can deplete nuclear NAD+, which then will stop SIRT1, SIRT6, and SIRT7 function. This then leads to a pseudohypoxic state of the nucleus due to HIF-1a stabilization. This then leads to inadequate TFAM, which then results in inadequate expression of mitochondrially encoded proteins for electron transport. Thus, mitochondrial dysfunction can be triggered by a telomere-dependent, DDR-mediated activation of p53, which affects nuclear encoded mitochondrial proteins. In addition, excessive DNA damage due to increased oxidative stress can deplete nuclear NAD, due to the requirements of PARP-1 for NAD as a cofactor. As a result, there is inadequate NAD for SIRT1, SIRT6, and SIRT7 function. This nuclear depletion of NAD then creates a pseudohypoxic nuclear state and a decrease in the expression of mitochondrial encoded proteins.
So, starting with telomere damage as the center it is possible to get to dysfunction of the mitochondria via a nuclear effect, and then the Warburg effect in the cytoplasm, which we know can lead to cancers and other pathologies which can lead to further telomere shortening. There is no “center” in such a process, only points of reference which lead to other points of reference. And many cofactors, state conditions and possible interventions can empower or retard such a process.
The villain factor here seems to be P53. In the final section of this blog entry, we argue that P53 is also the hero for conveying health and strength in the face of stresses.
Further, the catalytic component of telomerase hTERT can migrate out of the nucleus and if mutated create various forms of cell damage and dysfunction including in the mitochondria. This is another pathway through which a telomere-related problem can propagate into other cell components including mitochondria.
The 2010 publication A mutant telomerase defective in nuclear-cytoplasmic shuttling fails to immortalize cells and is associated with mitochondrial dysfunction reports: “Telomerase is a reverse transcriptase specialized in telomere synthesis. The enzyme is primarily nuclear where it elongates telomeres, but many reports show that the catalytic component of telomerase (in humans called hTERT) also localizes outside of the nucleus, including in mitochondria. Shuttling of hTERT between nucleus and cytoplasm and vice versa has been reported, and different proteins shown to regulate such translocation. Exactly why telomerase moves between subcellular compartments is still unclear. In this study we report that mutations that disrupt the nuclear export signal (NES) of hTERT render it nuclear but unable to immortalize cells despite retention of catalytic activity in vitro. Overexpression of the mutant protein in primary fibroblasts is associated with telomere-based cellular senescence, multinucleated cells and the activation of the DNA damage response genes ATM, Chk2 and p53. Mitochondria function is also impaired in the cells. We find that cells expressing the mutant hTERT produce high levels of mitochondrial reactive oxygen species and have damage in telomeric and extratelomeric DNA. Dysfunctional mitochondria are also observed in an ALT (alternative lengthening of telomeres) cell line that is insensitive to growth arrest induced by the mutant hTERT showing that mitochondrial impairment is not a consequence of the growth arrest. Our data indicate that mutations involving the NES of hTERT are associated with defects in telomere maintenance, mitochondrial function and cellular growth, and suggest targeting this region of hTERT as a potential new strategy for cancer treatment.”
Another important 2011 article relating telomere dysfunction to profound mitochondrial problems via activation of p53 and repression of PGC-1α and PGC-1β is Telomere dysfunction induces metabolic and mitochondrial compromise“Telomere dysfunction activates p53-mediated cellular growth arrest, senescence and apoptosis to drive progressive atrophy and functional decline in high-turnover tissues. The broader adverse impact of telomere dysfunction across many tissues including more quiescent systems prompted transcriptomic network analyses to identify common mechanisms operative in haematopoietic stem cells, heart and liver. These unbiased studies revealed profound repression of peroxisome proliferator-activated receptor gamma, coactivator 1 alpha and beta (PGC-1α and PGC-1β, also known as Ppargc1a and Ppargc1b, respectively) and the downstream network in mice null for either telomerase reverse transcriptase (Tert) or telomerase RNA component (Terc) genes. Consistent with PGCs as master regulators of mitochondrial physiology and metabolism, telomere dysfunction is associated with impaired mitochondrial biogenesis and function, decreased gluconeogenesis, cardiomyopathy, and increased reactive oxygen species. In the setting of telomere dysfunction, enforced Tert or PGC-1α expression or germline deletion of p53 (also known as Trp53) substantially restores PGC network expression, mitochondrial respiration, cardiac function and gluconeogenesis. We demonstrate that telomere dysfunction activates p53 which in turn binds and represses PGC-1α and PGC-1β promoters, thereby forging a direct link between telomere and mitochondrial biology. We propose that this telomere–p53–PGC axis contributes to organ and metabolic failure and to diminishing organismal fitness in the setting of telomere dysfunction.”
The DNA-mitochondrial Axis of Aging
Also, this 2012 article characterizes DNA damage and mitochondrial decline as the “Axis of aging:” Axis of ageing: telomeres, p53 and mitochondria. “Progressive DNA damage and mitochondrial decline are both considered to be prime instigators of natural ageing. Traditionally, these two pathways have been viewed largely in isolation. However, recent studies have revealed a molecular circuit that directly links DNA damage to compromised mitochondrial biogenesis and function via p53. This axis of ageing may account for both organ decline and disease development associated with advanced age and could illuminate a path for the development of relevant therapeutics.”
These articles portray a direct link between telomere dysfunction and mitochondrial dysfunction, perhaps explaining why many old people are so cold-sensitive and tired much of the time. Recapitulating the steps of what happens:
Telomere dysfunction =>
DNA damage response =>
p53 levels go up =>
High p53 inhibits the expression of both PGC-1a and PGC-1b
Genes for nuclear encoded mitochondrial proteins will NOT be expressed, as a result
Exercise cannot undo this and increase mitochondrial function.
Only deleting p53 or restoring telomere length will do this. This can be done with an Ornish/Blackburn lifestyle-type modification program
Telomeres, and why continuous, uninterrupted DNA damage response (DDR) signaling via p53 is so bad!
The reason is that you can’t turn such constitutive signaling off! Here is a 2012 article wrapping this up:Telomeres are favoured targets of a persistent DNA damage response in ageing and stress-induced senescence. “Telomeres are specialized nucleoprotein structures, which protect chromosome ends and have been implicated in the ageing process. Telomere shortening has been shown to contribute to a persistent DNA damage response (DDR) during replicative senescence, the irreversible loss of division potential of somatic cells. Similarly, persistent DDR foci can be found in stress-induced senescence, although their nature is not understood. Here we show, using immuno-fluorescent in situ hybridization and ChIP, that up to half of the DNA damage foci in stress-induced senescence are located at telomeres irrespective of telomerase activity. Moreover, live-cell imaging experiments reveal that all persistent foci are associated with telomeres. Finally, we report an age-dependent increase in frequencies of telomere-associated foci in gut and liver of mice, occurring irrespectively of telomere length. We conclude that telomeres are important targets for stress in vitro and in vivo and this has important consequences for the ageing process.”
By this point you should be getting the impression that P53 is a big-time villain when it comes to aging-related telomere attrition and damage that shows up in mitochondria and other systems. So be this. However, in the final section of this blog entry, we argue that P53 is also the hero for conveying health and strength in the face of stresses.
Again, in summary, including why telomerase-activating supplements cannot stop the axis of aging:
1. Live single cell studies showed that 50% of the DDR foci are located at telomeres (I suspect that the rest are associated with aneuploidy-related foci (such as the pericentromeric region) the nucleolus (due to rDNA), and possibly repetitive noncoding DNA.
2. Telomere-localized DDR foci occurs even if telomerase is activated (see article). This is why telomerase activating supplements such as Astralagus-based products do NOT turn off the DDR. (I.e. TA-65 or Product B). This is a very important point!
3. Telomere-localized DDR foci occurs in mouse liver and gut cells regardless of telomere length. This convinces me that the DDR signal from telomeres is NOT due primarily to telomere shortening or a lack of telomerase, but is due to a more complex “failure of telomere maintenance” that may be due to lack of SIRT6 activity, either due to a lack of nuclear NAD+ or a lack of SIRT6 gene expression secondary to SIRT6. Remember that the promoter site for SIRT6 has a binding site for SIRT1 (I.e. SIRT1 functions as a transcription factor for SIRT6 gene expression.
So there we have it. a. Yes, short telomeres can create all kinds of problems via DDR signaling, b. Problems due to telomere DDR signaling can also happen when the telomeres are long, c. Telomerase mutations can create serious mitochondrial problems, c. Telomerase=promoter extender supplements cannot solve the problems, and d. We need to look at interactions such as involving NAD+ and SIRT1 and SIRT6 if we want to positively affect the “Axis of aging.”
17. The catalylitic subunit of telomerase, telomerase reverse transcriptase (TERT) exercises a number of functions independent of telomere elongation.
The 2012 publication Telomere-independent functions of telomerase in nuclei, cytoplasm, and mitochondria reports: “Telomerase canonical activity at telomeres prevents telomere shortening, allowing chromosome stability and cellular proliferation. To perform this task, the catalytic subunit (telomerase reverse transcriptase, TERT) of the enzyme works as a reverse transcriptase together with the telomerase RNA component (TERC), adding telomeric repeats to DNA molecule ends. Growing evidence indicates that, besides the telomeric-DNA synthesis activity, TERT has additional functions in tumor development and is involved in many different biological processes, among which cellular proliferation, gene expression regulation, and mitochondrial functionality. TERT has been shown to act independently of TERC in the Wnt-β-catenin signaling pathway, regulating the expression of Wnt target genes, which play a role in development and tumorigenesis. Moreover, TERT RNA-dependent RNA polymerase activity has been found, leading to the genesis of double-stranded RNAs that act as precursor of silencing RNAs. In mitochondria, a TERT TERC-independent reverse transcriptase activity has been described that could play a role in the protection of mitochondrial integrity. In this review, we will discuss some of the extra-telomeric functions of telomerase.” (Italics ours)
The TERT subunit of telomerase and NF-κB p65 appear to be capable of up-regulating each other, at least in cancer cell lines. In fact, this could be an important positive loop involved in oncogenic transformation.
Theup-regulation of TERT by NF-kappaB appears to be mediated by c-Myc.
Nuclear factor-{kappa}B-mediated regulation of telomerase: the Myc link.
The 2013 publication Human telomerase reverse transcriptase regulates MMP expression independently of telomerase activity via NF-κB-dependent transcriptionreports: “Telomerase plays a pivotal role in the pathology of aging and cancer by controlling telomere length and integrity. However, accumulating evidence indicates that telomerase reverse transcriptase may have fundamental biological functions independent of its enzymatic activity in telomere maintenance. In this study, the ectopic expression of human telomerase reverse transcriptase (hTERT) and its catalytic mutant hTERT K626A induced cancer cell invasion accompanied by the up-regulation of the metalloproteinases (MMPs) MMP1, -3, -9, and -10. Both hTERT and hTERT K626A induced MMP9 mRNA expression and promoter activity in an NF-κB-dependent manner. hTERT and hTERT K626A also regulated the expression of several NF-κB target genes in cancer cell lines. Furthermore, both hTERT and hTERT K626A interacted with NF-κB p65 and increased NF-κB p65 nuclear accumulation and DNA binding. A mammalian 1-hybrid assay showed a functional interplay between hTERT and NF-κB p65 that may mediate NF-κB-dependent transcription activation in cells. Together, these data reveal a telomere-independent role for telomerase as a transcriptional modulator of the NF-κB signaling pathway and a possible contributor to cancer development and progression.”
The 2013 publication Transcriptional activation of hTERT through the NF-kappaB pathway in HTLV-I-transformed cellsreports: “In immortal cells, the existence of a mechanism for the maintenance of telomere length is critical. In most cases this is achieved by the reactivation of telomerase, a cellular reverse transcriptase that prevents telomere shortening. Here we report that the telomerase gene (hTERT) promoter is up-regulated during transmission of human T-cell lymphotropic virus type-I (HTLV-I) to primary T cells in vitro and in ex vivo adult T-cell leukemia/lymphoma (ATLL) samples, but not asymptomatic carriers. Although Tax impaired induction of human telomerase reverse transcriptase (hTERT) mRNA in response to mitogenic stimulation, transduction of Tax into primary lymphocytes was sufficient to activate and maintain telomerase expression and telomere length when cultured in the absence of any exogenous stimulation. Transient transfection assays revealed that Tax stimulates the hTERT promoter through the nuclear factor kappaB (NF-kappaB) pathway. Consistently, Tax mutants inactive for NF-kappaB activation could not activate the hTERT or sustain telomere length in transduced primary lymphocytes. Analysis of the hTERT promoter occupancy in vivo using chromatin immunoprecipitation assays suggested that an increased binding of c-Myc and Sp1 is involved in the NF-kappaB-mediated activation of the hTERT promoter. This study establishes the role of Tax in regulation of telomerase expression, which may cooperate with other functions of Tax to promote HTLV-I-associated adult T-cell leukemia.”
Cross-talk between sirtuins
As a final point, we can’t leave the six sister sirtuins of SIRT6 out of the telomere discussion. The sirtuins are far from independent actors in-vivo, exhibiting much cross talk among themselves as well as with numerous other factors. From Mammalian Sirtuins and Energy Metabolism (2011):“As one of the most important cellular metabolic sensors in cells, it has been speculated that the seven sirtuins coordinate with each other in various cellular compartments to actively monitor diverse environmental signals, modulating cellular metabolic activity, gene transcription, and genome stability, ultimately affecting aging. Indeed, perturbation of the activity of one sirtuin has been shown to impact the activities of other sirtuin members. For example, we reported that in mouse macrophages, deletion of SIRT1 leads to increased levels of chromatin-associated SIRT6 near the NF-κB binding sites, resulting in reduction of local acetyl-H3K9 levels, compensating for the hyperactivation of NF-κB induced by SIRT1 deficiency (119). Interestingly, this compensatory effect between SIRT1 and SIRT6 does not appear to exist in hepatocytes. Instead, deletion of SIRT1 causes 50% reduction of both SIRT6 mRNA and protein levels in the liver (97). This is a result of SIRT1 binding to Foxo3a and nuclear respiratory factor 1 (NRF-1) on the promoter of SIRT6 in hepatocytes, directly promoting the expression of SIRT6 under both basal and fasting conditions (97). Additional studies will be necessary to address whether the varying coordination patterns between these two nuclear sirtuins are related to the different metabolic profiles of macrophages and hepatocytes. — Intensive crosstalk between nuclear and mitochondrial sirtuins have also been implied in the literature. A recent study reported that SIRT3 is a transcriptional target of PGC-1α via an estrogen-related receptor binding element (ERRE) on its promoter (120). As PGC-1α is a direct deacetylation substrate of SIRT1 (42), this observation suggests that SIRT1 could indirectly modulate the expression of SIRT3 through PGC-1α deacetylation. Additionally, Nasrin et al. recently showed that SIRT4 knockdown in hepatocytes induces an increase in fatty acid oxidation through SIRT1 (116). Although molecular mechanisms underlying these connections remain to be defined, these findings suggest the existence of a sirtuin-network that may be pivotal in the maintenance of systemic metabolic homeostasis(ref).”
Towards a GUT, looked at from the end of the chromosome
Vince comment: We focus here on stress-response pathways, key to an emerging GUT in biology as pointed out in previous blog entries, particularly in the November 2013 Prospectus for a Grand Unified theory of Biology, Health and Aging. We have often argued in this blog that stress-response pathways offer numerous opportunities for generating and enhancing human health and possibly longevity via hormesis. But there has been an interesting paradox, which is that very many very different stresses can be used for the same end health objective, say preconditioning to prevent complications from a kind of surgery. I just attended a meeting of the International Dose-Response Society, where the focus was on stress-induced pre-conditioning(ref). I learned that among the stresses useful for surgical and other forms of preconditioning are exercise, gamma and x-radiation, pulses of laser light, pulsed DC current, heat, cold, restriction of blood circulation, calorie restriction, intermittent fasting, and hypoxia among others. It seems that a common denominator in hormesis is stress and that all these and many other stressors activate the same pathways. But the mechanisms through which this happens have been not known and has long been the subject of speculation. In the case of remote preconditioning, stress on one part of the body can lead to healthy impacts on other remote and seemingly unrelated parts of the body. In the conference I attended there was much discussion that there must be a “Factor X” which is the common mechanism for stress conditioning – an illusive factor capable of many pleiotropiceffects that marvelously work over a range of organisms and scale ranging from cells to organs to whole animals. It is possible that in the following Jim Watson has identified what this factor X is – simply expression of P53 which in small doses exercises beneficial effects and in large doses kills cells.
A good theory must be “summarizable” or “expressable” in a simple form. The following Summary is a descriptive form:
L = 2(GGAA). Where
L = Longevity
GGAA = transcription start site motif for all of the key genes involving DNA repair for both SS and DS breaks, genes involving the Shelterin proteins (Including Rap1), the hTERT gene, and the Genes affected by CR Mimetics. Here is the “story:”
Once upon a time, evolution decided that yeast and bacteria were not stress resistant enough to live longer than a few weeks. Evolution wanted to make organisms that had longer lifespans so evolution could blossom (I.e. provide a longer time for genetic diversity to evolve). So evolution decided to create a new, stress-responsive transcription start signal that would be activated by stress responses, such as running away from dinosaurs (exercise), surviving drought (starvation), blizzards (cold stress), and hot weather (the Ideal condition for evolution to flourish). The old TATA box method of activating gene transcription had too much “baggage”. The baggage was thousands of genes that were activated by “good times” such as lots of food (Insulin/IGF pathway), sex (hormones), and the lack of stress (I.e. the housekeeping genes and genes responsible for growth and development). So “Father Evolution” decided to make a new transcription start site for key genes that would be activated by stress. This way, only the most robust members of the species that were stress resistant would survive. For economy’s sake, evolution decided that this new transcription start site would be activated by the same transcription factor that would kill bad cells, such as Cancer cells. So Father Evolution decided to choose p53 as the transcription factor to bind to this new motif. After all, P53 being the Guardian of the Genome is the king of apoptosis when things in the cell are going too screwy. So why not use P53 for a different signaling function that would trigger very different actions in response to stress when the cell is functioning fairly normally? The Motif was GGAA duplicated and Papa Evolution put this right by the Start site for the binding of RNA polymerase II. He decided to put the genes responsive for DNA repair, Telomere stabilization, and epicentromeric stabilization all under the control of moderate, sublethal levels of P53 expression, below the 2nd NOEL point for p53. That is for levels of P53 in the hormetic range, below the dose level required to tall a cell to blow its brains out. (For every stress response there are two NOEL [No Observable Effect Level] points on the stress scale. For stress level between zero and the 1st NOEL point, the impact on the organism is negative. Stress levels between the 1st and 2nd NOEL points, in the hormetic range result in stress responses such that the net benefit to the organism is positive. Stress levels above the 2nd NOEL point result in a negative impact on the organism.)
Thus, evolution created the genetic equivalent of what The German philosopher Nietzsche said: “What does Not kill you (I.e. P53) makes you stronger (I.e. activated P53).”
Resveratrol and 2DG are both caloric restriction mimetics which Induce promoter activities of 5′ flanking regions of genes encoding telomere maintenance factors, including Shelterin proteins and hTERT. This occurs in both normal cells and cancer cells.
The protein, p53 is turned on by resveratrol and 2DG via AMPK phosphorylation, which then activates genes via the MAPK kinase ERK/2.
The P53 gene (TP53) also has a duplicated GGAA motif on the 5′ side of its transcription start site.
76% of transcription start sites in the human genome have no TATA box or TATA like elements.
(“The TATA box (also called Goldberg-Hogness box)[1] is a DNA sequence (cis-regulatory element) found in the promoter region of genes in archaea and eukaryotes;[2] approximately 24% of human genes contain a TATA box within the core promoter.[3] == Considered to be the core promoter sequence, it is the binding site of either general transcription factors or histones (the binding of a transcription factor blocks the binding of a histone and vice versa) and is involved in the process of transcription by RNA polymerase. == The TATA box has the core DNA sequence 5′-TATAAA-3′ or a variant, which is usually followed by three or more adenine bases. It is usually located 25 base pairs upstream of the transcription start site.(ref Wikipedia))”
There are 174 different binding motifs at the transcription start sites in human genes. TATA box or TATA like elements are one of the 174.
A large number of genes that maintain telomere health and DNA repair have a duplicated GGAA motif near their 5′ end of the transcription start site.
The protein p53 can bind to this duplicated GGAA motif. This may be the fundamental mechanism of how many different cellular stressors such as radiation, UV light, calorie restriction, etc induce life span or health span increases via resistance to cellular stress. Specifically, this may be the “Hormetic pathway” of gene activation for genes involved with DNA repair and telomere maintenance.
The next equation for the GUT must be the mathematical description of the biphasic dose response curve. The X-axis will be “dose” and the Y axis will be p53 gene activation via the double GGAA motif binding site at the 5′ end of genes that do the following:
Repair DNA – BRCA1, BRCA2, ATR, WRN, ATM
Control the cell cycle – RB, CDKN, etc.
Stabilize the end of the chromosome – Shelterin proteins, hTERT, subtelomeric DNA
Stabilize the center of the chromosome – I.e. Centromere and Pericentromeric DNA.
The mathematical formula must include 2 NOEL points which can be moved to the left or right based on the following:
FOXO3a-mediated gene expression (antioxidant and apoptosis factor levels)
NF-kB mediated gene expression
Etc
These factors would alter how much stress (I.e oxidative stress, radiation stress, toxin stress) we can handle before we pass NOEL point 2. There are likely to be many other factors that could push the NOEL points to the Left or right, such as NAD levels, AMP levels, Acetyl-CoA levels, SAM levels, alpha- ketoglutarate levels, and FAD levels. Thus the simple formula could have one “Placeholder” constant where each of these variables can be Inserted. Thus, we keep the formula “Clean” and Simple
Recapitulation and Summary, re. the GUT:
Once upon a time, a new switch for turning on genes was created by evolution to ensure fitness of the larger species that needed to live longer for biodiversity in evolution to occur. This new “start signal (GGAA x 2) was unique from the old switches (TATA) that turned on genes in response to plentiful times, such as growth factors, hormones, and plentiful food. This new “switch” would respond to stressors that conferred “fitness” to the species, allowing them to survive famine, radiation, ultraviolet light, drought, deserts, winters, and infections.
The genes that this new switch (GGAAGGAA) would need to turn on would keep DNA repaired, keep the telomeres stable, and keep the centromeres stable. Otherwise the organisms would have a short lifespan like yeast and bacteria. The “trigger” for this new “switch” would also have to be able to trigger cell death (apoptosis) if the levels of cellular damage became too high (Ex: Too much DNA damage, telomeres too short, or aneuploidy). For this reason, evolution decided to make p53 the trigger for both the GGAAGGAA motif at low levels of stress, whereas with high levels of stress, the classical DNA binding motif for p53 would be activated.
This Classical DNA binding motif for apoptosis or cell cycle arrest is as follows: (RRRCWWGYYY
Where R = A/G
W = A/T
Y = C/TThis motif can be separated by an optional spacer of additional base pairs to form a full binding site.
This Classical p53 binding motif is found at all transcription start sites for genes involving either p53-mediated apoptosis or p53 mediated cell cycle arrest (cellular senescence). To activate apoptosis, p53 must be acetylated at lysine 120 (K120) by the protein acetylase, Tip60.
K120 acetylation activates p53 binding to only apoptosis genes, not to cell cycle arrest genes (I.e. Cellular senescence). Thus, apoptosis can be selectively inhibited by deacetylation of K120 on p53 without turning off p53 mediated cell cycle arrest pathway.
Thus, p53 at low doses confers stress resistance. At higher doses, p53 induces apoptosis or cell cycle arrest. The cellular decision to undergo apoptosis vs cell cycle arrest is made by the cell and not an extrinsic factor. This decision depends on the acetylation status of K120. SIRT1 or SIRT2 deacetylate K120 and therefore induce cell survival, but will not necessarily inhibit cell cycle arrest, since this can occur with a deacetylated p53. These are the two Janusian faces of P53.
So, the “axis of fitness” and the “axis of aging” mentioned above are both due to P53 expression. Which one will be choosen depends on a decision in the cell which depends on dose and the state of the cell.
This is the story of life.
On good vs bad
The more I learn, I now believe that ROS can be good or bad – it depends on the dose, where the ROS is being made (mitochondrial source vs plasma membrane NOX), and the levels of anti-oxidant enzymes produced by both Nrf2 and FoxO3a pathways.
The same for cellular senescence – it depends on what cells are undergoing senescence (white blood cells in wound healing, precancerous cells being shut off, or cells undergoing aging), where they are located (i.e. chronic inflammation), and how much the STAT3-dependent SASP factors are being expressed (i.e. the SASPs, like IL-6, IL-8, etc.)
The same for apoptosis – it depends on what cells are undergoing apoptosis (cancer cells vs normal embryonic development vs hippocampal neurons, etc.). It also depends on whether replacement cells are being made from stem cells (Ex: skin, colon, muscle, brain, etc. ).
The same for p53 – it depends on the dose (low confers resistance to stress, high confers either apoptosis or cellular senescence), and whether p53 is acetylated at K120 or not (i.e. apoptosis vs cell cycle arrest).
The same for SIRT – it depends on whether the cell is normal or malignant (with normal cells, activating SIRT1 is good, with cancer cells, it could be bad).
Frequent readers of this blog are familiar with the fundamental importance that we have attributed to biological stresses and stress-responses in driving health and longevity. The two of us attended the 2014 meeting of The professional society that is concerned with such stresses, the International Dose Response Society, held on the campus of the University of Massachusetts in Amherst Massachusetts April.22-23. The conference was organized by Dr. Edward J. Calabrese, a pioneer and continuing contributor in the field of adaptive biological stress responses. The theme of the Conference this year was Preconditioning – Adaptive Responses in Biology and Medicine – Building Biological Shields Against Disease and Injury. The purpose of this blog entry is to review and comment on some of the presentations at that meeting that we think have particular significance. We selectively quote from Conference abstracts.
GENERAL FRAMEWORK
What is preconditioning?
In the broadest context, preconditioning is the use of stress to yield an improved health outcome or set of outcomes. The best known example is exercise which of course yields multiple benefits. Another example is fasting or alternate-day fasting (ref) which also induces many health benefits including a number of reduced disease susceptibilities. In a narrower medical context, preconditioning is the application of controlled stress to mitigate against the anticipated negative effect of from an anticipated later stress, or to produce a better specific health outcome. Examples include: 1.Remote Ischemic/hypoxic conditioning: Inducing ischemia for brief periods with an arm pressure cuff to protect against ischemic reperfusion injury resulting from subsequent heart surgery, or to protect against brain injury due to stroke. 2 Use of low dose radiation to protect against diabetes-induced renal damage, and 3. Use of heat or cold or hypoxic stress to protect against brain injury that could be incurred in a boxing match or football game, or as an unintended consequence of brain surgery.
Preconditioning has long been a topic of both research and practice in medicine. Specific preconditioning techniques appear throughout the history of medicine. An example is the use of moxibustion in ancient as well as contemporary Chinese folk medicine(ref). A search on the term “preconditioning: in Pubmed.org reveals 11,590 research publication citations going back to 1949, and many additional citations not in the database hark back to earlier dates. Indeed, the idea of remote preconditioning goes back to 1933.
Preconditioning is the practical application of Hormesis
Early in the Conference, Ed Calabrese (Environmental Health Sciences, School of Public Health, University of Massachusetts Amherst) , pointed out that preconditioning is a central application of hormesis, a universal adaptive stress-response phenomenon of all biological systems. We have often written about hormesis in this blog. (See for example the PowerPoint presentation on Multifactorial Hormesis which can be downloaded by going to this link). We have also argued that hormesis is an essential component of an emerging Grand Unified Theory of biology, health and aging(ref).
The 2008 publication Hormesis Defined by Mark P. Mattson (one of the key and initial speakers at the Conference), relates: “Hormesis is a term used by toxicologists to refer to a biphasic dose response to an environmental agent characterized by a low dose stimulation or beneficial effect and a high dose inhibitory or toxic effect. In the fields of biology and medicine hormesis is defined as an adaptive response of cells and organisms to a moderate (usually intermittent) stress. Examples include ischemic preconditioning, exercise, dietary energy restriction and exposures to low doses of certain phytochemicals. Recent findings have elucidated the cellular signaling pathways and molecular mechanisms that mediate hormetic responses which typically involve enzymes such as kinases and deacetylases, and transcription factors such as Nrf-2 and NF-κB. As a result, cells increase their production of cytoprotective and restorative proteins including growth factors, phase 2 and antioxidant enzymes, and protein chaperones. A better understanding of hormesis mechanisms at the cellular and molecular levels is leading to novel approaches for the prevention and treatment of many different diseases.”
Here is the abstract of Ed Calabrese’s opening remarks relating preconditioning to hormesis: “Optimizing Pre- and Post-conditioning Clinical Outcomes: A Dose Response Perspective: “This study assessed the dosage, temporal and mechanistic relationships between the conditioning dose and the protective effects of preconditioning experiments. Entry criteria for study evaluation required the occurrence of an hormetic-like biphasic dose response for the protective endpoint, and a mechanistic assessment of how the conditioning dose affected the protective endpoint response. The conditioning dose that demonstrated the largest increase in endpoint response during the conditioning period was the same dose that was the optimally protective dose. Cell signaling pathway inhibitors were commonly employed to block the conditioning effect; such inhibitory actions abolished the protective effect at the optimal conditioning dose, identifying a specific hormetic mechanism. Conditioning dose responses often had sufficient doses to assess the nature of the dose response. In each of these cases these mechanism-based endpoints displayed an hormetic dose response. The present analysis reveals that when preconditioning experiments demonstrate a biphasic dose response it can be directly linked to the actions of the conditioning dose based on optimal dosage, temporal relationship and receptor-based and/or cell signaling-based mechanisms. These findings indicate that preconditioning induced biological/biomedical effects represent a specific type of hormetic dose response.”
The cogency of relating preconditioning to hormesis is very important because it appears that many researchers have worked for years in specific areas of preconditioning focusing on specific interventions for very specific objectives like ischemic conditioning prior to cardiac surgery. Some of these researchers have done important research but have never paid attention to the term “hormesis” and are not familiar with the general universality of stress-response mechanisms in biology. As we have pointed out in multiple blog entries, hormesis applies to essentially all biological systems, on all levels of scale (e.g. cell, organ and whole organism), relates to all the stresses an organism ordinarily encounters, is essential for functioning of any organism from a basic control-systems perspective, and is important for understanding the stress triggers of evolution(ref)(ref)(ref).
In the course of the Conference, there was also discussion of post-conditioning –applying a stress after a traumatic event such as a head injury or operation, and perconditioning, simultaneous conditioning during a stress event, such as applying a stress during an operation. In general, all of these seem to produce beneficial responses.
Conference content
The PDF book of abstracts for the Conference can be downloaded by clicking 2014_Abstract_book. In this blog entry, we will cover and comment on only selected presentations and emphasize what we believe to be central themes of importance. We do this within our own framework of organization.
Two of the most important stressors for inducing general health and neurological health in particular are our old familiar friends – exercise and fasting.
One of the first presentations in the opening general session was by Mark P. Mattson, Laboratory of Neurosciences, National Institute on Aging Intramural Research Program, and entitled Intermittent Energetic Challenges, Adaptive Responses and Health: Lessons from the Brain. “Humans evolved in environments where food was not available ad libitum, and so possess robust adaptive physiological and behavioral responses to periods of food scarcity. Emerging research in this Laboratory and elsewhere has shown that intermittent fasting (IF; e.g., fasting for a period of 24 hours twice weekly) and vigorous exercise can increase numbers and strength of synapses and can enhance brain function (cognitive and sensory – motor performance) and mood. We find that the general mechanism by which IF and exercise benefit neurons is by challenging them by increasing their activation state and energy demand, which results in a coordinated engagement of signaling pathways that promote neuroplasticity and cellular stress resistance. The pathways activated by exercise and IF include those involving brain-derived neurotrophic factor (BDNF), mitochondrial biogenesis, DNA repair and removal of oxidatively damaged proteins and organelles. Peripheral changes in energy metabolism that occur during fasting and exercise may also contribute to their beneficial effects on the brain. In this regard, the depletion of glycogen stores in the liver triggers the mobilization of fatty acids from fat cells and the production of ketone bodies. Ketone bodies such as beta-hydroxybutyrate provide an alternative energy source for neurons and may also activate signaling pathways that enhance the ability of the brain to cope with stress. Our studies in animal models of chronic neurodegenerative disorders (Alzheimer’s and Parkinson’s diseases) and acute brain injury (stroke and severe epileptic seizures) demonstrate robust neuroprotective and neurorestorative effects of IF diets. IF protects the brain by bolstering antioxidant defenses and protein chaperone levels, and by suppressing inflammation. The implications of these findings for strategies for optimizing brain function and reducing the risk of neurodegenerative disorders will be described.”
Note a common theme here that applies to hormesis and preconditioning in general: A hormetic stress response can and usually does involve simultaneous upgrading of multiple pathways producing multiple health benefits for the whole organism. This is very unlike the kind of response typically sought by pharmaceutical company researchers – which is to find a particular kinase drug which upgrades one particular biological pathway to achieve a specific health objective. Thus, hormesis is not subject to a basic limitation of this drug-oriented approach. Because of the multiplicity of interacting pathways which exist in-vivo, a kinase drug approach which works in test tubes may not work or produce the opposite effect in-vivo. This, we believe, is a basic reason why so many clinical trials of seemingly good drug candidates fail.
An excellent example of preconditioning researched over a period of more than 80 years is remote ischemic preconditioning
The abstract for the Conference presentation Remote Ischemic Conditioning: From Inspiration to Clinical Translationby Karin Przyklenk of the Cardiovascular Research Institute and Departments of Physiology & Emergency Medicine, Wayne State University School of Medicine is: “Remote ischemic conditioning is the phenomenon whereby brief episodes of ischemia-reperfusion applied in a distant organ or tissue render the heart resistant to damage caused by a prolonged ischemic stress. The discovery of remote conditioning, first reported by our laboratory in 1993, was not a serendipitous finding, but, rather, was predicted by mathematical modeling. In the ensuing years, the paradigm has been expanded to encompass a spectrum of remote triggers, including the seminal observation that repeated 5 min periods of limb ischemia, achieved by inflation-deflation of a blood pressure cuff, initiated a profound cardioprotective response. The hallmark of remote ischemic conditioning – i.e., reduction of myocardial infarct size – has been documented and confirmed in multiple models and species. Two major, remaining challenges are to elucidate the cellular and molecular mechanisms responsible for this intriguing form of cardioprotection, and successfully exploit the infarct-sparing effect of remote ischemic conditioning for the treatment of patients with acute myocardial infarction.”
Some common themes identified here are 1. Conditioning can be remote, it does not have to be a procedure exercised on the target organ to be effective, 2. Conditioning can be initiated by a variety of different stress triggers, 3. Conditioning can be via a very simple and easy to administer stress such as by employing a blood pressure cuff. Remote ischemic preconditioning has been the subject of at least 24 randomized clinical trials related to a multiplicity of clinical situations including abdominal aortic aneurysm repair, open heart surgery, percutaneous coronary intervention, living donor renal transplantation, coronary angiography, elective decompression surgery, carotid endarterectomy, recent stroke, or transient ischemic attack combined with intracranial carotid artery stenosis(ref).
Conditioning can provide long-lasting additional benefits beyond immediate protection against event-induced injury
Another presentation on ischemic preconditioning was by Andrew N Redington, Division of Cardiology, Department of Paediatric Cardiology, Hospital for Sick Children, University of Toronto entitled Assessing the Promise of Remote Conditioning for Cardioprotection: New Clinical Developments. “Remote ischemic conditioning (RIC) is a powerful, innate, mechanism of protection against ischemia-reperfusion injury. This simple, non-invasive stimulus can be induced in patients by 3-4 cycles of 5-10 minutes inflation (ischemia) and deflation (reperfusion) of a standard blood pressure cuff, placed on a limb. Following our original description of the method in 2002, and our ‘first-in-human’ RCT showing reduced markers of cardiac damage, and improved myocardial and lung function in children receiving RIC prior to open heart surgery, there have been multiple adult cardiac surgical trials. Most, but not all studies have shown benefit, with recent meta-analyses showing an overall benefit in terms of biomarkers of injury, and in a recently published 5-year follow-up study of one of the largest studies showing early benefit, those randomized to RIC, had a lower rate of adverse events and lower mortality compared to the controls. Other proof-of-principle RCT’s show that RIC; reduces myocardial injury and renal failure in abdominal aortic aneurysm surgery; reduces incidence of renal dysfunction due to contrast-induced nephropathy; improves early outcomes in elective percutaneous coronary intervention (PCI); and our own study by Botker et al showing improved myocardial salvage overall, and reduced infarct size, when RIC is administered prior to emergency PCI. The late outcome data from these studies are also emerging. In a follow-up of the CRISP study (RIC in elective PCI) there was a significant reduction in major adverse cardiovascular events (MACE) at 6 years, and both MACE and mortality rates were reduced at 5 year follow up of Botkers original study of RIC in emergency PCI for evolving myocardial infarction (MI). Finally, early experimental data suggests that repeated, daily, RIC may have additional benefits on post-MI remodeling, and a single RCT suggests that daily RIC improves stroke recovery.”
Here we encounter additional general properties of remote conditioning, again reflecting that conditioning is an application of hormesis: 1. Although the stress may have been applied to protect against specific damage due to a specific event like a pending surgery, unexpected benefits or forms of protection may show up (e.g. increased lung capacity for a stress induced to protect against ischemia-reperfusion injury) 2. The beneficial results may be very long lasting or permanent, 3 . Application of the stress can lead to reduction in mortality, and 4. Daily or periodic repetition of the stress can lead to additional benefits. Indeed, 5. conditioning for one purpose in one system can possibly produce benefits in other body systems, e.g. ischemic pre-surgery conditioning can possibly produce neurological benefits such as greater capability to focus and perform cognitive tasks, better body balance, etc.
The cardioprotective effects of exercise appear to be associated with an enhanced capability to buffer ROS stress, and that the protective adaptive response is triggered by exercise-induced ROS
This point was made in the Conference presentation by David A. Brown, Department of Physiology, Brody School of Medicine, East Carolina University,Cellular Mechanisms Underlying the Cardioprotective Effects of Exercise: “Numerous pre-clinical and epidemiological studies have documented the cardioprotective efficacy of exercise, yet the mechanisms that underlie exercise-induced preconditioning of the heart are not fully understood. During conditions when reactive oxygen species (ROS) overwhelm endogenous buffering systems, the loss of mitochondrial function is closely linked with the onset of cardiac electromechanical dysfunction. In a series of studies using both short- and long-term treadmill-running protocols, we have established that exercise preconditions the rat heart in the absence of markers of systemic stress. Fluorescence studies in cardiac myocytes from exercised rats indicated that endogenous ROS buffering-capacity was augmented, and correlated with a lower propensity for cell death. During cardiac ischemia-reperfusion, glutathione levels in exercised hearts were better maintained in Langendorff-perfused hearts, which was associated with lower arrhythmia scores and smaller infarct sizes. While resting myocardial glutathione levels were not different between sedentary and exercised animals, the replenishment of glutathione was enhanced in exercised hearts, due to up-regulated glutathione reductase enzyme activity. The beneficial effects of exercise were abolished when NADPH oxidase activity was pharmacologically inhibited during exercise, but remained intact when mitochondrial ROS production was blunted during exercise. These data lead us to believe that ROS produced during exercise ‘trigger’ adaptive responses, and that the source of ROS does not appear to be mitochondrial in origin. Our most recent studies have directly examined mitochondrial energetics in the intact heart using multi-photon microscopy, and preliminary data indicate that the preservation of mitochondrial membrane potential was directly linked to prevention of reperfusion arrhythmia. In conclusion, our studies provide novel insight into adaptive responses following exercise that protects the heart during acute coronary syndromes.”
Protection against damage induced by stroke is another benefit of ischemic conditioning, an objective not achieved by recognized pharmacological approaches.
This topic was discussed in a Conference presentation Ischemic Tolerance and Neurological Protection was offered by John M. Hallenbeck, Chief, Stroke Branch, NINDS, NIH. “Ischemic tolerance can be interpreted as a form of hormesis in which a stress that is gauged to be sub-lethal can activate evolutionarily conserved, endogenous protective mechanisms and induce a time-limited tolerance to an otherwise damaging or lethal stress. There are various forms of induced tolerance to ischemic stress that have been modeled preclinically. They include preconditioning in which the sublethal stress is applied shortly before the otherwise damaging ischemia (immediate preconditioning) or that precedes the damaging ischemia by 24-72 hours (delayed preconditioning). Additional forms of induced tolerance include perconditioning (stress superimposed during damaging ischemia) , post-conditioning (stress imposed after damaging ischemia), remote conditioning (another organ or tissue site than brain is exposed to ischemic stress that then induces tolerance to damaging brain ischemia), and cross-conditioning (a non-ischemic stress induces tolerance to ischemia). The stroke field is broadly interested in developing an understanding of the molecular mechanisms that regulate tolerance in order to help guide efforts to develop cytoprotective therapies for strokes. This interest is understandable when one realizes that vascular neurology research has yet to translate a cytoprotective therapy that has been identified by standard reductionist techniques and have it show efficacy in a Phase III clinical stroke trial.”
Conditioning for stroke was also discussed in the Conference presentation Post-conditioning and the Transition from Animal Models to Humans for the Treatment of Stroke by
Roger Simon, MD, Neuroscience Institute, of the Morehouse School of Medicine “Endogenous mechanisms of protection against stroke and acute injury can be demonstrated in brain and other organs. Such therapies might replace or enhance putative pharmacotherapy. The induction of endogenous protection is via a response to sub lethal stress which induces “preconditioning”. The preconditioned organ is then “tolerant” to injury from subsequent severe stress of the same or different etiology (ie brief ischemic stress protects against injury from prolonged epileptic seizures and vice versa). Ischemic preconditioning provides protection against subsequent stroke. Protection is substantial (70% reduction) but delayed and transient (onset at two days, maximum at three days and gone by seven days). Gene expression is unique between brains preconditioned, injured (stroke) or made tolerant. Thus, preconditioning reprograms the brains response to lethal stress (stroke); reprogrammed from an injury induction response to a neuroprotective processes. Transcriptional down regulation is the central feature of the reprogrammed response to injury. This process of neuroprotective gene silencing is driven epigenetically via polycomb group proteins which suppress transcription broadly. — Postconditioning refers to attenuation of injurious processes which occur during reperfusion of ischemic brain (the only approved treatment for acute stroke). Mechanically inturupting referfusion induces postconditioning which can attenuate reperfusion injury. Postconditioning protects ischemic brain by decreasing reperfusion induced oxygen free radical formation. The free radicals produce injury via mitochondrial damage which can be repaired experimentally with resultant neuroprotection as potent as experimental postconditioning. The recognition of broad based gene silencing (suppression of thousands of genes) as the phenotype of the ischemic tolerant brain, may explain the failure of all single target drugs for stroke. The risks of reperfusion treatment for stroke may be attenuated by induction of endogenous repair processes. Thus endogenous neuroprotective and repair mechanisms offer translational stroke therapy.”
We underlined the final point here because we believe it identifies a very important distinction between hormetic stress interventions and most pharmacological interventions. If we want to realize significant objectives like protecting ourselves against stroke or heart damage or if we want to create other significant health benefits, in most cases that involves upregulation or down regulation of hundreds or thousands of genes. This happens naturally through mobilization of our endogenous stress response pathways but can’t be done using a single protein kinase.
Another general property of preconditioning and hormesis appears to be that repeated intermittent periods of stress can build a stronger and longer lasting stress response capability than that achievable by a single application of the stress.
There seems to be much folk knowledge and evidence that this is true when exercise and fasting are adapted as regular practices. An interesting question is the extent to which it is true for other types of stress like ischemic conditioning. The presentation Promoting Long-lasting Protection in the CNSdiscusses repeated hypoxic stress.It is by Jeffrey M Gidday, of the Department of Neurosurgery, Department of Ophthalmology & Visual Sciences, and Department of Cell Biology & Physiology, Washington University School of Medicine. “Significant reductions in the extent of acute injury in the CNS can be achieved by exposure to a preconditioning stimulus, but the duration of the protective phenotype is short-lasting. Our work has been directed at extending the period over which such epigenetic changes persist, thereby enhancing translational relevance. Having established in adult mice that a single exposure to systemic hypoxia (2 h of 11% oxygen) provides transient (a few days) protection against cerebral (Miller BA et al., NeuroReport, 2000) and retinal (Zhu Y et al., IOVS, 2002) ischemia, we then documented that repetitive presentations of this same hypoxic stimulus over a two-week period (six total exposures to 2-h of 11% oxygen, every other day over two wks) extended the duration of ischemic tolerance to one month after the last preconditioning stimulus (Zhu Y et al., IOVS, 2007). Protection against stroke for two months could be afforded by a 2-wk intermittent hypoxic preconditioning regimen (Stowe AM et al., Annals Neurol, 2011), but only with stochastic increases in stimulus frequency, duration, and intensity, suggestive of tissue- or cell-dependent hormetic dose-response relationships. Given this protracted ‘therapeutic window’, we then sought to determine whether repetitive hypoxic conditioning (RHC) could also enhance cell survival in chronic neurodegenerative disease. In an inducible mouse model of glaucoma defined by progressive loss of retinal ganglion cell soma and axons over a 10-wk period of elevated intraocular pressure (IOP), significant protection of both soma and axons could be demonstrated − without a reduction in IOP − when animals completed the 2-wk RHC treatment before IOP elevation (Zhu Y et al., Mol Med, 2012), or received it during the period of elevated IOP (manuscript in preparation). Thus, extending the duration of the adaptive epigenetic phenotype by RHC represents a fundamentally new therapeutic approach for treating glaucoma and other chronic neurodegenerative diseases.”
With regard to hypoxia. we have frequently heard skeptics say things like “It is not worth holding your breath if you are waiting for a big breakthrough in health and longevity.” We think there is a better basis for saying “Holding your breath may provide you a small breakthrough in health and longevity.”
Many different stresses can be used for preconditioning for multiple health objectives
The sheer numbers of stresses and health benefits may appear amazing, that is until we remember that stress conditioning is applied hormesis and that a biphasic stress response probably applies to every kind of stress that biological organisms and their subsystems have encountered and adapted to in their evolutionary histories. These presentations discussed different stimuli for conditioning for different objectives:
Pre-conditioning with Low Level Laser (Light) Therapy Tanupriya Agrawal, MD, Wellman Center for Photomedicine at Massachusetts General Hospital; James D Carroll, FRSM, Thor Photomedicine Ltd; Michael R Hamblin, Ph.D., Wellman Center for Photomedicine at Massachusetts General Hospital
Dose-Response Effects of Low-Level Light Therapy on Brain and MuscleF. Gonzalez-Lima, Departments of Psychology, Pharmacology and Toxicology, University of Texas at Austin,
Neuromodulation with Weak Transcranial Electrical Stimulation: Small Things Making a Big Difference Marom Bikson, Neural Engineering Lab, Department of Biomedical Engineering, The City College of New York of CUNY,
Increased Threat Detection, Learning and Attention using Low-Level Transcranial Direct Current Stimulation (tDCS) Vincent P. Clark, Psychology Clinical Neuroscience Center, University of New Mexico and MIND Research Network, Dept. Psychology, University of New Mexico.
There are multiple examples of preconditioning that can provide health benefits to us humans where we are not the actual targets of the preconditioning.
A presentation at the Conference discussed stress-conditioning stem cells to increase their survivability, vitality and potency prior to transplantation into humans. The presentation Preconditioning Strategy for Improved Therapeutic Potential of Stem Cell Transplantation Therapy after Ischemic Stroke was by Shan Ping Yu, Emory University School of Medicine, Atlanta. “Ischemic stroke is a serious threat to human life and health but clinical treatment for stroke is very limited. Stem cell transplantation has emerged as a promising regenerative medicine for stroke and neurodegenerative disorders. It is expected the repair of damaged brain tissues/structures can be achieved using pluripotent/multipotent stem cells derived from embryos, fetuses, or even adult tissues. However, many issues and problems remain to be resolved before successful clinical applications of the cell-based therapy. Among them, poor cell survival, uncertain neuronal differentiation and low efficacy of tissue repair in the harsh microenvironment of the injured brain are some primary issues. We have initiated an effort to develop a combination strategy in stem cell transplantation therapy in order to improve the therapeutic potential of transplanted cells. One of the major focuses has sought to benefit from well-known mechanisms of ischemic/hypoxic preconditioning that activates cellular defense mechanisms and shows marked protective effects against multiple insults found in ischemic stroke and other acute attacks. A sub-lethal hypoxic exposure significantly increases the expression of pro-survival and pro-regenerative factors. So far, a variety of preconditioning triggers have been tested on different stem cells and progenitor cells. Preconditioned cells show much better cell survival, increased neuronal differentiation, and enhanced paracrine effects that lead to increased trophic support and tissue repair. Transplantation of preconditioned cells helps to suppress inflammatory factors and immune responses. The combination therapy also includes strategies for increasing migration and homing of transplanted cells to the lesion site. More importantly, the combination stem cell therapy shows much improved morphological repair as well as functional recovery after ischemic stroke. Although the combination strategy in stem cell transplantation is still an emerging research area, accumulating information from reports over the last few years already indicates it as an attractive, if not essential, prerequisite for transplanted cells.”
We have been concerned with another category of non-human preconditioning that can benefit humans: stress-preconditioning fruits and vegetables to lead them to produce and contain enhanced amounts of health-producing phytochemicals when we eat them. We have discussed such interventions in this blog(ref)(ref)(ref).
One topic of central concern in the Conference was why the adoption of preconditioning in standard medical procedures has been so slow to happen even though the benefits have been known for a great many years. This applies both to preconditioning for specific purposes, such as protection against adverse consequences of surgery, and for creating a more general health state and longevity.
One example is that the benefits of calorie restriction for health and longevity in mammals has been researched extensively since 1934, but that calorie restriction practices such as intermittent fasting have not been adopted in mainline medicine. In the Conference, Mark Mattson commented on this situation and what might be done about it in his presentation Implementation of Intermittent Fasting Prescriptions: Breaking Through the Barriers: “Compelling evidence from studies of rodents and humans suggest that intermittent fasting (IF), consisting of periodic (1-4 days per week) short (16-36 hours) fasts, can promote optimal health and can prevent and reverse disease processes in many chronic conditions including diabetes, cancers, cardiovascular disease and neurodegenerative brain disorders. In overweight human subjects, IF diets promote long-term weight loss with retention of lean mass, increased insulin sensitivity, and reduced inflammation and oxidative stress. Why, despite the fact that IF is a safe and effective intervention, do health care providers not prescribe IF diets to their patients? Regrettably, the reason for this lack of effort by the medical community is that no one profits from IF prescriptions. The processed food and agriculture industries would lose money if people ate less, and the pharmaceutical industry would ‘suffer’ if fewer people developed the diseases for which Pharma peddles their drugs. Medical training and practice is focused on technologically advanced treatments and specialists (cardiologists, neurologist, orthopedists, etc.) that it is not their job to tackle the underlying cause (which is often a couch potato lifestyle) of their patients’ diseases; instead, they have become drug-dispensing, scalpel-wielding robots. Medical school curricula are devoid of training on the profound health benefits of IF and exercise, and primary physicians often assume that patients will not comply with IF diets. In this presentation I will describe strategies for the implementation of IF diets in which patients are given a specific plan for the diet and for monitoring their progress. The physician and/or an assistant is in close communication with the patient via text messaging, social media, etc. with the purpose of guiding them through the 1 – 2 month period that is often required for a person to adjust to the IF eating pattern.”
Poster Presentations
As is usually the case at the annual International Dose Response Society Meetings, there were a number of quite interesting poster presentations dealing with related topics. Many of these are summarized in the abstract book. The titles are:
Astrocyte Preconditioning by Severe Stress is Glutathione- but not Heat Shock Protein 70-Dependent
Drosophila melanogaster Show a Threshold Effect in Response to Radiation
How Radiotherapy Was Historically Used to Treat Pneumonia: Could it be Useful Today?
Use of X-rays to Treat Shoulder Tendonitis/Bursitis: A Historical Assessment
Model Uncertainty in Cancer Risk Assessment
Risk Assessment Report Card
Low-dose radiation Prevents Diabetic Complications Protection of Hearts Against Ischemic Insult: Changes in Iron Homeostasis Explain Myocardial Response to Preconditioning
Investigation of the Cellular and Molecular Mechanisms of Radiation-induced Bystander Effects in a Human Keratinocyte Cell Line
Low-level radiation and heart disease death rates in four states: An ecological study Inter-Relationships between Low Dose Hypersensitivity, Induced Radioresistance and Bystander Effects in Human Cell Lines
Comparative Acute Toxicity of Silver Nanoparticles Produced by Physical (Top-Down) and Chemical (Bottom-Up) Methods in Zebrafish (Danio rerio)
Ultraviolet-A photoemission from cells upon β-irradiation and consequent bystander effects
Physiological Conditioning Hormesis Improves Post-Irradiation Performance in Young and Aging Fruit Flies
Communication of Protective Signals from Fish Sub-Lethally Challenged with Vibrio anguillarum VIB1 to Naïve Fish
Assessing Predictive Factors and Radiation-Induced Non-Targeted Effects in Blood Serum from Cancer Patients
The Effects of Chronic Exposure to Low Levels of Alpha-Emitting Radionuclides on the Health and Reproductive Fitness of Mammals
Pre-operative Stress Conditioning: Role for Hyperbaric Oxygen Therapy
Drosphila melanogaster Show a Threshold Effect in Response to Radiation
Protection of Hearts against Ischemic Insult: Changes in Iron Homeostasis Explain Myocardial Response to Preconditioning
Correcting deficiencies in our societal structure for the Application of Science in Medicine
The final wrapup session
The final session of the conference was devoted to a Integrative Discussion Of How Can Pre-Post Conditioning Being Used. Among the topics treated there were therapeutic applications, protecting military personnel, preventing athletic injuries and enhancing recovering, enhancing athletic and job performance, and improving public health. Again, a major focus of the discussion was inquiring why penetration of conditioning techniques into mainline medicine has been so painfully slow despite the fact that the efficacy and safety of such techniques have been known for many years. A number of barriers were identified, including ones referred to above in Mark Mattson’s presentation, ones related to the current structuring of medical practice and the reward patterns in that field, and ones related to the nature of the pharmaceutical industry. One of the barriers identified was the continuing absence of identifying a common molecular mechanism that could allow so very many different stress stimuli to produce similar or identical effects. In the discussion this unknown mechanism was called Factor X. We point out that Jim Watson has suggested a strong candidate for Factor X in the previous blog entry Nuclear Aging: The View from the Telomere end of the Chromosome, the Section near the end entitled Towards a GUT, looked at from the end of the chromosome suggests that Factor X is none other but our old molecular friend P53 which serves as a hormetic signaling molecule at low doses and a death signaling (apoptotic) molecule at high doses.
This is a progress report on the changing state of human longevity during the five-year lifespan of this blog. It reviews major trends and forces impacting on our lifespans, both scientific and social. Longevity can be viewed either from the societal viewpoint or the personal viewpoint. It is impossible to comprehend one fully without considering the other. From a personal viewpoint, enhanced longevity may come from taking a supplement or a “longevity drug” or pursuing a certain lifestyle practice. And it also can come about as a result of background happenings in our society such as public health measures: cleaner air and water, availability of bike and walking paths, and less tobacco smoke. I review some important trends here that are manifest today, and discuss how I see them to be interacting so as to create longer and healthier lives. Five years ago, I speculated that the efficacy of practical interventions for life extension can be expected to double every seven years, proposing this as “Giuliano’s Law.” At the end of this blog entry I look again at that speculation.
So, this is a “big picture” blog entry looking mainly at the interaction of multiple factors impacting on longevity, how they relate, and how they are unfolding. It is a progress report, a snapshot from today’s view. The main content of this blog is a summary review of trends with links to many (but by far not all) of the some 460 blog entries we have generated.
TRENDS
Jim Watson and I have published blogs of “Top-12 things I have learned about aging in 2013(ref)(ref),” This blog goes further back and offers a five-year view, one that combines objective developments and personal opinion.
1. SOCIETIES ARE AGING. As a general trend, in all healthy societies in the world we are living longer and longer. This is an observed and unmistakable phenomenon, a trend since before the beginning of history that continues to accelerate. The trend is widely recognized by international agencies and has resulted in new commercial initiatives to address the needs of aging and retiring “baby boomers.” We have written about it extensively(ref)(ref)(ref)(ref).
Here is one graphical look at the history of human lifespans and projecting where they are headed:
Image source, The line for the United States in 1970 has moved to the right to where it is now close to the leftmost dotted line, The authors of the source article project human longevity increase in humans based on animal studies if calorie restriction mimetics can be found which effectively control inflammation and inflammatory-related diseases. I am quite sure that achieving radical life extension will require addressing many additional biological pathways in addition to inflammatory ones, but that the projected extension of human lifespans may well be possible.
2. SCIENCE OF AGING KEEPS EXPANDING: There have been significant advances in scientific understanding of fundamental biological processes and how they impact on aging
One field hardly visible to me five years ago but now of major proportions is epigenitics. DNA Methylation, histone acetylation and chromatin remodeling are the classical areas of epigenetics. The new frontier in epigenetics appears to be understanding the roles of the multiple species of non-coding RNAs and understanding what particular RNAs impact on gene regulation in which particular circumstances. Of particular interest are long non-coding RNAs, micro RNAs, and other RNA species such as circular RNAs. We have written several blog entries about these topics(ref)(ref)(ref)(ref)..
Of great interest also is what happens to DNA and RNAs with aging. Concern includes age-related global and local DNA methylation, proliferation of long repeat DNA sequences and circular RNAs, and changes in the DNA repair machinery as it is impacted by various molecular processes and aging(ref)(ref)(ref)(ref)(ref)(ref)(ref)
Another area where there has been an immense swelling of research during the last five years has to do with molecular pathways that can regress cells to earlier age states, particularly all the research related to induced pluripotent stem cells (iPSCs). There is a burgeoning field of research in regenerative medicine involving the use of iPSCs as more and more stem cell therapies are coming into actual use (ref)(ref)(ref)(ref)(ref)(ref).
The period has seen deepening of understanding as to the functioning of key molecular pathways related to life and aging and how these pathways interact. We see that we can start with any key biological system in the cell whether this is the nucleus, the mitochondria chromosome telomeres or other organelles, and trace interacting pathways to essentially all other important biological systems and organelles. One of several ways to start looking at aging, for example, is in terms of REDOX states(ref). Another perspective is identified in the blog entries on the view from the end of the chromosome(ref)(ref)(ref). Autophagy is another key process related to both health and longevity(ref). Also, we can start with any key longevity related molecule like SIRT1, IGF-I, Nrf2 or Klotho and trace how these impact on key longevity pathways which in turn impact on multiple other important pathways. In summary, we are continuing to discover the complete systems nature of biology which consists of numerous interacting control and homeostasis–maintaining loops. (ref)(ref)(ref)(ref)(ref)(ref)(ref).
3. NEW DISCOVERIES CAN COME FROM THEORY OR OBSERVATIONS. Observed longevity phenomena are leading to theoretical understanding as well as the other way around. These in turn inform interventions which can enhance health spans and lifespans. Not all science starts with a theory and works towards practical results. In many cases an observation leads to a whole body of theoretical understanding. This is the case, for example for the mTOR pathway, where suppression of this pathway is a proven way to extend longevity in a number of animal models. mTOR stands for mammalian target of rapamycin a substance that was discovered in the soil of Easter Island and later refined into the drug rapamycin, So first came the observation of what rapamycin did and after that came understanding of the mTOR pathway which is suppressed by rapamycin. Examples of discoveries during the last five years and being very much discussed today include:
Heterochronic parabiosis and GDF 11. It was observed that when the bloodstream of an older mouse is crossed with that of a paired younger mouse, the older mouse experiences rejuvenation in numerous dimensions, while the younger mouse appears to get older. This has led to a search for the factor or factors in the blood involved in this rejuvenation. One of the factors has been recently discovered, GDF 11. This factor is known to produce rejuvenation effects in the heart, cardiovascular system, brain and nervous system and possibly other systems as well. Some key publications related to the discoveries appeared only this month(ref)(ref)(ref)(ref)(ref).
Mitochondria. There have been a series of research discoveries associated with mitochondrial health, On the one hand, these discoveries involve understandings of key molecular biological pathways, such as how events in a cell nucleus can impact what goes on in mitochondria, how dysfunctions in telomere-maintenance pathways can lead to problems in DNA repair and mitochondrial respiration and redox state. Also there is increased understanding about the possibility of practical interventions to enhance mitochondrial health and possibly longevity: NMM, NR, mitoQ, and buckyballs (ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref).
Diseases. There has been increasing understanding of the processes involved in age-related disease processes, not only cancers (ref)(ref)(ref)(ref)(ref)(ref) but also many others including Parkinson’s disease(ref), sarcopenia(ref), MRSA(ref), lymphoma(ref)(ref), lupus(ref), and infectious diseases(ref).
Interspecies interdependencies. Another area of increasing both theoretical and practical interest relates to bacterial biomes, interspecies signaling, quorum sensing and high degree of interspecies interdependency(ref)(ref)(ref). There has been particular interest in the gut biome, for example(ref).
Circadian rhythms, These profoundly affect and are affected by gene expression(ref)(ref)(ref).
Neural and brain health and neurogenesis, and muscle health, all in relationship to aging (ref)(ref)(ref)(ref).
Pathway links between inflammation, aging and cancers, There may be interventions that serve to minimize all three of these(ref)(ref)(ref))(ref)(ref).
Key molecules. We have often returned to certain molecules, particularly ones that serve key signaling functions, including P53, NF-kappaB, IGF1, species of ROS, sirtuins, Nrf2, heat and cold-shock proteins, and key gasses(ref)(ref)(ref)(ref)(ref).
Some findings have been negative. For example, telomere extending supplements are unlikely to work(ref)(ref) and you can forget about 1-a-day multivitamin supplements(ref). The old free radical theory of aging is dead(ref) and is replaced with a much more sophisticated version involving mitochondria.
Systems biology and quantum biology offer new and alternative perspectives on biological phenomena(ref)(ref),
Klotho-related results. There has been a steady increase in the perceived importance of Klotho for longevity, mental health, and retaining full cognitive capability in aging. For background, you can check out my 2011 review article on Klotho. There have been many important subsequent results and I plan to publish an updated blog entry on the subject soon. In the interim, you may want to check out a few relevant research publications(ref)(ref)(ref)(ref)(ref).
Key cell components. We have looked at additional key cell components beyond those in the nucleus and mitochondria, and the roles they play in health maintenance disease processes and aging. An example is microtubules(ref). Another is the endoplasmic reticulum(ref)(ref).
There is exponential growth in the sciences related to aging. Publication rate provides a measure. For example, consider items indexed in PUBMED, the general medical-related literature:
PMID publications in PUBMED
YEAR NUMBER
ENTRIES
NUMBER ADDED
% INCREASE
2009:
17.764,826
2010:
18.502,916
738,090
4.15
2011:
19,569,568
1,066,653
5.76
2912
20,494,848
925,280
4.73
2013
21,508,439
1,013,591
4.95
So, the cumulative impact for seven years would be around 45% and growth is more or less exponential. However the growth rates for medium-hot area of research related to longevity are much greater. For example, for sirtuin, there were 16,286 publications in 2006 and seven years later there were 69,936, an increase of 329% For klotho there were 43 publications in 2006 and 192 in 2013, an increase of 347%. For iPSC there were 47 publications in 2006 and 434 in 2013, an increase of 823%. For ncRNA there were 47 publications in 2006 and 168 in 2013, an increase of 257%. These numbers suggest far more than doubling of knowledge in key longevity-related topics over a 7 year period.
Grand Unified Theory of biology, Health and Aging (GUT). As we have been studying more and more biological pathways and phenonena and integrating our knowledge gleaned from diverse disciplines, I believe the outlines of a comprehensive GUT are starting to emerge, as well as new insights into the process of evolution (ref)(ref)(ref)(ref). I believe Jim Watson joins me in this perception.
APPLIED SCIENCE:
The last five years have, for me at least, seen the emergence of a number of new and practical health and antiaging interventions, ones soundly grounded in research and that can be widely adopted today. These interventions can involve approaches such as stress pathway activation to generate hormetic stress signaling, overcoming age-related restraints on gene activation such as DNA promoter site methylation, and promoting mitochondrial health.
Uses of specific phytosubstances to promote health via specific and sometimes multiple pathways, neuroprotection and protection against cancer, microglial activation and other disease processes. Exploitation of xenohormetic substances and plant-based RNAs. (ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref).
Practical applications of hormesis and conditioning approaches as well as improved understanding of stress-response pathways and how to use them to promote health(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref)(ref).
Uses of liposomal and nano-formulations of healthful substances to greatly enhance their bioavailability and increase effective doses without incurring side effects(ref)(ref)(ref).
There has been a stream of new findings relating to health properties of food substances. Also, we have published original articles in this blog related to stressing fruits and vegetables to increase their healthfulness. (ref)(ref)(ref)(ref).
Longitudinal observations of centenarians and super-centenarian population cohorts have also been yielding a wealth of observations relating to genetics, lifestyles, diets and the stresses encountered(ref)(ref)(ref)(ref)(ref). The results of these observational studies can be correlated with molecular biology studies of centenarians(ref). In my opinion, they lend credibility to suggested practical interventions we have identified for longevity such as use of hormetic stresses as well as careful attention to diet.
In summary with respect to science, there has been an increasing rate of breakthrough discoveries and upwards integration of knowledge. We have already identified and are increasingly testing novel longevity-enhancing strategies. And we may be getting closer to discovering even more-powerful anti-aging interventions
There have been steady improvements in engineering technologies related to research, biotechnology and practices. These include better and lower cost diagnostic tools and techniques, more and better predictive biomarkers, more economic abilities to do large scale scanning, extension beyond genome scanning to include epigenetic indicators, methylome’s, transcriptomes, improved bioreactors and other bioproduct manufacturing techniques and equipment, etc, Genetic Engineering and Biotechnology News is a definitive biotechnology sector trade publication which announces a variety of new approaches, innovative and lower-cost biotech technology every month.
Big Biotech is a powerful economic engine propelling life sciences innovation. The world biotech industry had sales of $47 billion in 2006 and estimated $133 billion in 2013. A 7-year increase of 183%. This industry thrives on innovation. I believe it plays a similar role in the 21st century life-sciences revolution that the semiconductor industry played in the 20th century information and communications revolution.
“Big Data” and bioinformatic tools are allowing a variety of studies leading to levels of understanding hitherto impossible. One application is determining biological pathways common to multiple disease processes as well as aging(ref); another is to accelerate drug discovery(ref). Another application is targeted repurposing of already-approved drugs and their use in personalized medicine(ref). There are additional applications of big data for personalized medicine(ref) and for improving the effectiveness of healthcare(ref).
There is increasing consciousness on the part of the population of the importance of maintaining wellness(ref)
new campaigns focusing on diet, particularly with respect to obesity and type 2 diabetes
increasing public sophistication with regard to diets
acceleration of focus on fitness
new focus on disease prevention in the Affordable Care Act(ref)
Personal electronic activity and biomarker tracking. More small sophisticated wearable devices, “smart scales” and other personal health-indicator measuring devices are coming on the market, ones that constantly or daily monitor health indicators and communicate them wirelessly to a central personal “health and fitness” database. Marketed as health and fitness devices instead of medical devices, they are available to all consumers. The market for these devices is becoming highly competitive, there being a large number of “activity trackers” now being sold. Fitbit sells tiny gadgets that measure steps, flights of stairs climbed, estimates calories burned daily and tracks sleep patterns. Using the fitbit Aria scale, the measurements also include weight, body lean-to-fat ratio and BMI. Performance charts and records are automatically uploaded and available on computers and mobile devices anywhere. The Withings devices measure the same indicators plus pulse rate, blood pressure and blood oxygen. I expect we will see more and more biomarkers included in the list as time goes on, a relatively easy picking being blood glucose for example. I expect there will be evolution to the point where hundreds of health biomarkers will be automatically tracked and sophisticated software will provide interpretations, daily health guidance and medical alerts.
Health and wellness apps. The Apple iTunes Store (iOS) and Google Play (Android) each offers hundreds of Health and Fitness apps(ref)(ref)(ref) for smart cellphones and tablet devices. Most are free or cost less than $4. In 2009, these did not exist. Last week there was news that Apple is focusing on health and fitness in iOS 8 with its new ‘Healthbook’ App(ref).
The maturation of the Baby Boomer population. Large numbers of people are entering the range where they are susceptible to age-related diseases but wish to maintain their own health and wellness. The size and affluence of the Boomer market provides a major incentive for entrepreneurial innovation related to health and wellness. Social innovations may have major health impacts. Examples include bike lanes in cities, subsidized exercise and wellness services, and public health campaigns. A large number of commercial organizations are mobilizing to support the Boomer population and have increased their focus to the affluent retiring boomer population. Traditional organizations concerned with elders, like the AARP, have shifted their focus to the health wellness and vitality of those over 50.(ref)(ref)(ref)(ref).
Innovations in wellness and health care are supplementing the existing highly-institutionalized health care system. There is ongoing expansion in the numbers and scopes of employee wellness programs(ref). AARP just sponsored a LivePitch event in Boston on May 9, 2014, where innovators-entrepreneurs pitched their health innovations for people 50 and over. Videos of the pitches can be linked to here.
Walking and biking for health. Many progressive city and local governments are building bike lanes and paths, closing streets to auto traffic, enhancing public transportation options, implementing rail-to-trail paths and taking other steps that encourage walking and biking exercise while reducing auto emissions(ref).
Shifts in medical practice paradigms. These are evolving slowly but definitely towards being more science-based. For example, there is closer tying of cancer therapies to biomarker indicators of specific kinds of cancers. This is part of a long-term shift to personalize health and medicine(ref)(ref).
New public health initiatives like ones to further curtail smoking, to control childhood obesity by getting harmful sugars, starches and fats out of school lunch menus, and for getting kids moving. There is greater focus on air and water quality, automobile safety, product safety, workplace safety, etc.(ref)
Market for healthier foods. There is slowly growing appreciation for basic rules of nutrition. Along with this, immense markets have emerged for organically grown fruits and vegetables, free-range meats, cage-free chickens, and low-calorie and low fat products. Supermarket shelves feature increasing numbers of products which claim to be “all-natural” “fat-free,” “sugar-free” or “gluten free.” In many cases, such health claims associated with packaged foods are misleading or irrelevant. For example, a number of sugar-laden breakfast cereals feature on their boxes in giant print that they contain vitamins and minerals. Other products announce that they are “fat-free” but in fact contain large amounts of sugar and are hardly good for you. However, food labels are more stringent than they used to be, allowing aware consumers to discover packaged food contents if they take time to read the fine print.
Rules associated with medical reimbursement. Some changes in these mitigate towards a greater focus on health and wellness. An example is the Hospital Re-admissions Reduction Program, which reduces government payments to hospitals which have too many re-admissions. Thus, hospitals have a financial incentive to emphasize the importance of wellness measures to patients being discharged(ref). This is one example of how the runaway costs of the current healthcare system in the United States is creating powerful incentives for wellness. The results will be enhanced longevity
Wellness is emerging as a major thrust of large institutions who have played major roles in conventional health care delivery including health insurance companies and HMOs. Johnson and Johnson, United Healthcare and Kaiser Permenente are examples of such organizations.(ref)(ref)(ref)(ref).
IMPLICATIONS
From the broadest perspective, a combination of better scientific knowledge, social trends and initiatives, industry and engineering developments are already propelling the general populations in our country and in other advanced societies towards greater health and enhanced longevity. It is not just that science will do this in the future; it is already happening by the interaction of science, social and commercial developments and engineering developments. Extending human lifespans is not just something that is going to be in the future. It is something very much with us in the fabric of what is happening right now.
From a personal perspective, I believe that the swelling stream of scientific knowledge about health and longevity is increasingly enabling earlier adapters to live lives that are longer, healthier, and more productive than the lives experienced in the general population. Enhanced life extension is increasingly available for those willing to learn about how to pursue it and who are willing to modify their lifestyles and habits to bring it about.
Is such a projection reasonable? Many highly reputable scientists think so. “Earlier this year, researchers at the U.K.’s University of Cambridge and the Medical Research Council reported that people who exercise regularly, don’t smoke, limit their alcohol intake and eat five servings of fruit and vegetables a day live, on average, 14 years longer than people who didn’t.(ref)”
Reviewing Giuliano’s Law
How do I assess all the above in terms of effectiveness of practical longevity interventions? In March 2009, I posted a blog entry identifying what I called Giuliano’s Law (I must admit, a rather immodest naming act). As originally stated:
Starting now, every seven years will see the emergence of practical age-extension interventions (ones that have a potential of leading to extraordinary longevity) that double the power of the interventions available at the start of the 7 year period. That is, on an average basis, the practical anti-aging interventions available at the end of a seven-year period will enable twice the number of years of life extension than did the interventions available at the start of the period. Life extension is measured in years of life expectancy beyond those actuarially predicted for a given population starting in a certain base year.
I predicated Giuliano’s Law based on Moore’s Law as a model – the Law for microprocessor chips that stated: every two years, at a given price point, the power of a commercially available chip approximately doubles. That Law had been in operation for four decades in 2009. And it still appears to be operational today. For many of those years the doubling period was only 18 months, though I think it is now somewhat more than two years. I asked myself what were the scientific, engineering, commercial, economic and social factors that were driving Moore’s Law starting in the 1970s. The operation of the Law was and continues to be driven by a combination of such driving forces, not just on any one, but by a synergistic combination of all of them. And then I asked myself if there were equally compelling and interacting driving force factors affecting health and longevity today. The answer was definitely yes, and this was the basis on which I predicated Giuliano’s Law. I published a subsequent blog entries back in 2009: Factors that drive Giuliano’s Law which laid out the driving forces behind the operation of the Law. In a sense, this current blog entry is an update of that one, laying out driving forces for the Law as they appear even more powerfully today, five years later. And, several of the factors listed above were not visible in 2009. Thus, it may be that Giuliano’s Law with its projected 100% increase in effectiveness longevity interventions over a 7-year period is too conservative. Only time will tell.
Of course Giuliano’s Law has to be in the nature of a statistical projection. Time of death for anyone will be a stochastic variable depending on numerous personal and environmental factors and on luck. If you get mashed badly by a bus, a careful history of longevity interventions won’t help.
I strongly suspect that the Law is valid although 8 years may not be the correct doubling period My own anti-aging regimen has evolved considerably from what it was when I generated the Law five years ago and I am doing fine, but I will tell my personal story in a different blog entry..
Acknowledgments. The blog entries referred to here were generated by multiple associate researcher-writers as well as myself: Victor, Dan Campagnoli, Brendan Hussey, Melody Winnig and Mike Giuliano. Jim Watson has been a particularly important contributor for over a year now, not only for his original writings but also for our intense and stimulating phone and e-mail collaborative interactions. Melody Winnig has also served an importance literature surveillance function, bringing important findings to Jim’s and my attention on a daily basis, and as a blog proofreader.
In this blog, you must register as a subscriber in order to post comments. If you have had trouble registering, have found your registration cancelled or are having trouble posting comments, please read on.
I turned off new registrations for a few days last week because we were receiving many phony new registrations by spam bloggers (sploggers). These are people who use robots to register on multiple blogs and to post comments which advertise products or refer back to their own websites Sploggers do this in order to enhance the search engine rankings of their own sites and do not care about our blog or any other one they attack. A typical robot-generated comment by a splogger could read “Nice and interesting blog. You might want to check out my site at www.happyheartburn.com.” Another one from yesterday is “Welcome to visit my blog! you can find your dream.” Some spam comments are detailed boldface advertisements such as for leather boots or imported Viagra or products I don’t endorse like HGH.
For some time I have used software to block most such comments, but still the splogger registrations kept rolling in. And in the last few days several spam comments snuck through. Last night I ran a program that purged our subscriber list of known or suspected sploggers. And now I have turned the registration process back on. So, if you have mistakingly been thrown off the subscriber list, I invite you to re-subscribe.
I may have to implement a Captcha system where registering people have to figure out and copy weird letters.
And please if you are having trouble registering, posting comments or otherwise using this blog, let me know by writing me at at vegiuliano@agingsciences.com
By Vince Giuliano with important contributions by James P Watson
Image may be NSFW. Clik here to view.Glucosamine is one of the most popular dietary supplements, used by millions who hope that it will lessen the ravages of osteoarthritis, arthritis, loss of cartilage and associated pain and discomfort. This natural shellfish-derived substance has been known since 1876 and its biological impacts were reported as far back as in 1919(ref). I have suggested taking Glucosamine since the 2008 version of the treatise Antiaging Firewalls – The Science and Technology of Longevity.
Most of us have long thought of this substance as having a narrow purpose, focused on combating osteoarthritis and on cartilage regeneration . However, there has been mounting evidence that regular supplementation with glucosamine can significantly enhance longevity – in mammals probably as much as well studied interventions such as taking rapamycin. Here, I review studies that provide strong evidence for this prospect.
Research evidence for the new take on glucosamine is of several kinds: 1. biological mechanism of operation, 2. animal longevity impact studies and 3. human epidemiological studies . I start here with the first and second and then move to the third. Finally. I briefly review how the biological mechanisms of glucosamine compare to those of important alternative approaches to mitochondrial health.
If you are not familiar with metabolic processes in mitochondria and the concept of mitohormesis, you might want to review a few past blog entries as background to this one:
Biological activities of glucosamine and animal studies
The most recent and most powerful study supporting you longevity impacts of glucosamine appeared in April 2014: D-Glucosamine supplementation extends life span of nematodes and of ageing mice: “d-Glucosamine (GlcN) is a freely available and commonly used dietary supplement potentially promoting cartilage health in humans, which also acts as an inhibitor of glycolysis. Here we show that GlcN, independent of the hexosamine pathway, extendsCaenorhabditis eleganslife span by impairing glucose metabolism that activates AMP-activated protein kinase (AMPK/AAK-2) and increases mitochondrial biogenesis. Consistent with the concept of mitohormesis, GlcN promotes increased formation of mitochondrial reactive oxygen species (ROS) culminating in increased expression of the nematodalamino acid-transporter 1(aat-1) gene. Ameliorating mitochondrial ROS formation or impairment ofaat-1-expression abolishes GlcN-mediated life span extension in an NRF2/SKN-1-dependent fashion. Unlike other calorie restriction mimetics, such as 2-deoxyglucose, GlcN extends life span of ageing C57BL/6 mice, which show an induction of mitochondrial biogenesis, lowered blood glucose levels, enhanced expression of several murine amino-acid transporters, as well as increased amino-acid catabolism. Taken together, we provide evidence that GlcN extends life span in evolutionary distinct species by mimicking a low-carbohydrate diet.”
Image source. It appears that average lifespan extension for mice is about 8 weeks or 6%
From the article: “Discussion: The current findings indicate that GlcN at pharmacologically relevant concentrations is capable of extending life span in C. elegans and ageing mice. This appears to be a result of decreased glycolysis and a compensatory increase of amino-acid turnover. Although it should be noted that GlcN may impact hexosamine metabolism34, we did not, however, detect any differences in downstream metabolites of GlcN-6-phosphate in mice, nor did the disruption of the hexosamine pathway prevent GlcN from extending life span in worms, together strongly suggesting that hexosamine metabolism has no bearing on the phenotypes observed. Mechanistically, the life span-extending phenotype is rather linked to an activation of SKN-1/NRF-2-dependent transcription, as the latter not only promotes longevity of C. elegans independently of GlcN16,32,38,39,40,41,42 but also following GlcN exposure (this study). In this regard, it is interesting to note that the aat-1 gene contains a proximally located SKN-1-binding site and that GlcN exposure induces aat-1 ranscription through SKN-1. This notion is supported by the observations that impairment of either skn-1 or aat-1 abrogates the effects of GlcN on life span and disruption of SKN-1 prohibits the induction of aat-1 expression in nematodes. Interestingly, this upregulation of amino-acid transporters, as well as increased L-leucine/L-isoleucine metabolism, is found in GlcN-exposed liver specimen of rodents, thereby suggesting that the glycolytic inhibitor GlcN induces a metabolic switch towards increased protein metabolism in both worms and mice, unambiguously culminating in extended life span. – Although the specific cause(s) of death of individual mice have not been determined in the current study, it will be interesting to see whether and how GlcN affects not only glucose metabolism, but also cancer growth in chronically supplemented mice, as a number of studies insinuate that GlcN effectively reduces cancer cell proliferation43,44,45,46,47,48. — Unlike for DOG and most other life span-extending compounds, extensive published evidence indicates that GlcN is safe for human use even at high doses, making it readily available for interventions to extend human healthspan particularly because, on an observational and uncontrolled basis, it has been repeatedly suggested that supplementation with GlcN may decrease overall mortality in humans49,50.”
Here are some technical highlights from the paper as summarized by Jim Watson:
1. Glc-N: a Glycolysis inhibitor – The mechanism by which D-Glucosamine inhibits glycolysis is by inhibiting hexokinase (or the liver analog, glucokinase). This impairs glucose metabolism and results in the cell “switching” over to amino acid metabolism to produce energy. Thus D-Glucosamine is a “metabolic switch” that converts “Warburg-type metabolism” in the cell (i.e. aerobic glycolysis in the presence of adequate oxygen) over to “protein metabolism”. 100 microM of Glc-N results in a 43% inhibition of glucose oxidation. I don’t know if these levels are achievable with most oral Glc-N supplements.
On the other hand, 2-Deoxyglucose reduces glucose oxidation by over 90%. However. 2DG has been shown to reduce lifespan in rodents although it increases lifespan in nematoides. Glc-N does not have such a negative effect.
Glc-N: an AMPK activator - The mechanism by which glycolysis inhibition produces longevity is via AMPK(ref). AMPK is an “Energy sensing enzyme” that is activated when the cells energy levelsare low (I.e. Low ATP, high ADP and AMP). Glc-N therapy results in an ATP deficiency which then activates the regulatory unit of AMPK in the nematode, which is AAK-2. AAK-2 is activated with Glc-N treatment. AMPK is the main “on switch” for mitochondrial biogenesis. It turns on over 1500 nuclear encoded genes via a gene activating Cofactor called PGC-1a in the nucleus of the cell. Also, AMPK is a powerful autophagy activator.
3. Glc-N: mechanism is not significantly dependent on Sirtuins – They used the Glc-N on a Sir-2 deficient strain of nematodes as well. There was a reduced effect that did not change the lifespan of the nematodes in these Sir-2 deficient worms. This suggests that the mechanism of action of Glc-N is independent of nuclear levels of NAD+.
4. The mitochondrial biogenesis was due to AMPK - This is not surprising, since AMPK is the primary activator of PGC-1a. However, the article mentions that mitochondrial ROS generation transiently increased for 7 days after Glc-N treatment, but then normalized when the antioxidant enzymes MnSOD and catalase were unregulated.
This suggests that there is an increase in ROS-activated antioxidant gene expression, which probably is due to an Nrf2/Keap dependent mechanism. We have discussed NRF2/Keap extensively in this blog (ref)(ref)(ref).
5. Antioxidants prevent Glc-N-mediated lifespan extension - this is yet another added piece of evidence to the mounting pile of evidence that exogenous antioxidants can be bad for us. They used N-acetylcystein and butylated hydroxylanisole. I think this is an important “take-home message” that we must get out.
6. Mitohormesis – Transient Increase in ROS required for producing oxidative stress-resistant worms. This I believe is one of the most important lessons to learn from this study. Rarely do you see scientists outside the hormesis community who use the word “mitohormesis“. This is good to see. However, I must qualify this by saying that the “upstream issues” that produce the mitohormesis are sill glycolysis inhibition and AMPK activation. Mitohormesis is “downstream event” in this cascade, not the “upstream event”.
7. MAPK pathways and Nrf2 pathways are both important in mediating the lifespan lengthening effects of Glc-N. This again goes back to what we have discussed many times. Specifically, the p38/PMK-1 version of the MAPK pathway is activated with Glc-N treatment. This is a ROS-dependent activation. Likewise, the Nrf2 pathway activation is obviously ROS-dependent as well.
8. Glc-N intake in humans does not increase blood glucose -This is good news, but still puzzling. I would have expected that the inhibition of glycolysis would have raised blood glucose (via TORC2 inhibition)
9. SKN-1: the transcription factor that turns on 36% of the genes affected by GlcN.The paper mentions that when they analyzed promoter sites ofr GlcN-dependent genes, SKN-1 was the activator for 36% of the genes. We recall that this result is for nematodes and SKN-1 is the nematode counterpart of NRF2 in mammals.
10. N-acetyl-glucosamine has a different mechanism of action from D-glucosamine – The last paragraph in the discussion section of this paper mentions that another group has shown a lifespan extending effect of N-acetyl-glucosamine, but the mechanism of life span extension is by inhibiting the hexosamine pathway, not via AMPK.
11. Glc-N is safe, unlike 2-Deoxyglucose – 2DG has a lot of side effects. So does Metformin and Rapamycin. For this reason, GlcN may be a much better compound for humans for lifespan extension.
Glucosamine may also be the basis of anti-cancer therapies
One mechanism may be down-regulation of expression of the heat shock survival protein HIF-1α in cancer cells. The 2014 publication Short-term treatment with glucosamine hydrochloride specifically downregulates hypoxia-inducible factor-1α at the protein level in YD-8 human tongue cancer cellsreports: “Hypoxia-inducible factor-1 (HIF-1) is a tumor angiogenic transcription factor composed of an α and β subunit. We investigated the effect of glucosamine hydrochloride (GS-HCl) on the expression of HIF-1α and HIF-1β in serumtreated YD-8 human tongue cancer cells. While long-term (24 h) treatment with GS-HCl strongly repressed the expression of HIF-1α and HIF-1β at both the protein and mRNA levels, short-term (4 h) GS-HCl treatment inhibited HIF-1α at the protein level. Short-term GS-HCl treatment also decreased phosphorylation of p70S6K and S6, translation-related proteins. However, the results of subsequent pharmacological inhibition and protein stability analyses indicated that HIF-1α protein downregulation induced by short-term GS-HCl treatment was not through modulation of the mTOR/p70S6K/S6 signaling pathways, the 26S proteasomal and lysosomal activities and HIF-1α protein stability. Importantly, our further analyses identified that HIF-1α protein downregulation induced by short-term GS-HCl treatment was blunted by exogenous administration of the citric acid cycle metabolites citrate and 2-oxoglutarate, but not the glycolytic end byproducts pyruvate and lactate. These findings demonstrate firstly that short-term GS treatment selectively downregulates HIF-1α at the protein level in YD-8 cells via interference of production of the citric acid cycle metabolites. It is proposed that short-term GS-HCl exposure may be applied for the treatment of oral tumors with high expression of HIF-1α.”
Anti-cancer effects of glucosmine have been long-noted. The 1969 publication d-Glucosamine and Other Sugar Analogs on the Viability and Transplantability of Ascites Tumor Cells reported in summary: “d-Glucosamine has been shown to have a powerful cytotoxic effect on various ascites tumor lines, resulting in a decrease in viability and transplantability of the neoplastic cells. The toxic effect of glucosamine was not significantly altered by addition of either glucose or pyruvate. The effects of d-glucosamine and other sugar analogs were evident histologically even before the cells became stainable by trypan blue.”
The 2007 publication d-Glucosamine inhibits proliferation of human cancer cells through inhibition of p70S6Kreports: “Although d-glucosamine has been reported as an inhibitor of tumor growth both in vivo and in vitro, the mechanism for the anticancer effect of d-glucosamine is still unclear. Since there are several reports suggesting d-glucosamine inhibits protein synthesis, we examined whether d-glucosamine affects p70S6K activity, an important signaling molecule involved in protein translation. In the present study, we found d-glucosamine inhibited the activity of p70S6K and the proliferation of DU145 prostate cancer cells and MDA-MB-231 breast cancer cells. d-Glucosamine decreased phosphorylation of p70S6K, and its downstream substrates RPS6, and eIF-4B, but not mTOR and 4EBP1 in DU145 cells, suggesting that d-glucosamine induced inhibition of p70S6K is not through the inhibition of mTOR. In addition, d-glucosamine enhanced the growth inhibitory effects of rapamycin, a specific inhibitor of mTOR. These findings suggest that d-glucosamine can inhibit growth of cancer cells through dephosphorylation of p70S6K.”
Other publications relating glucosamine to cancers include:
Glucosamine induces cell death via autophagy in some cancer cell lines.
The research literature also indicates that glucosamine induces autophagy, a process of cellular house cleaning extremely important for health that we have discussed previously(ref)(ref). This is not surprising since AMPK is a powerful autophagy activator.
One is the 2010 article Glucosamine induces autophagic cell death through the stimulation of ER stress in human glioma cancer cells“Autophagy can promote cell survival or death, but the molecular basis of its dual role in cancer is not well understood. Here, we report that glucosamine induces autophagic cell death through the stimulation of endoplasmic reticulum (ER) stress in U87MG human glioma cancer cells. Treatment with glucosamine reduced cell viability and increased the expression of LC3 II and GFP-LC3 fluorescence puncta, which are indicative of autophagic cell death. The glucosamine-mediated suppression of cell viability was reversed by treatment with an autophagy inhibitor, 3-MA, and interfering RNA against Atg5. Glucosamine-induced ER stress was manifested by the induction of BiP, IRE1α, and phospho-eIF2α expression. Chemical chaperon 4-PBA reduced ER stress and thereby inhibited glucosamine-induced autophagic cell death. Taken together, our data suggest that glucosamine induces autophagic cell death by inducing ER stress in U87MG glioma cancer cells and provide new insight into the potential anticancer properties of glucosamine.”
Another relevant article just appeared a few days ago (June 2014) Enhanced expression of proapoptotic and autophagic proteins involved in the cell death of glioblastoma induced by synthetic glycans, Further this article suggests that glucosamine could possibly serve as a therapy for the deadly and incurable brain cancer glioblastoma, “OBJECT – Glioblastoma is the most aggressive malignant brain tumor, and overall patient survival has not been prolonged even by conventional therapies. Previously, the authors found that chemically synthesized glycans could be anticancer agents against growth of a series of cancer cells. In this study, the authors examined the effects of glycans on the growth of glioblastoma cells both in vitro and in vivo. METHODS – The authors investigated not only the occurrence of changes in the cell signaling molecules and expression levels of various proteins related to cell death, but also a mouse model involving the injection of glioblastoma cells following the administration of synthetic glycans. RESULTS – Synthetic glycans inhibited the growth of glioblastoma cells, induced the apoptosis of the cells with cleaved poly (adenosine diphosphate-ribose) polymerase (PARP) expression and DNA fragmentation, and also caused autophagy, as shown by the detection of autophagosome proteins and monodansylcadaverine staining. Furthermore, tumor growth in the in vivo mouse model was significantly inhibited. A dramatic induction of programmed cell death was found in glioblastoma cells after treatment with synthetic glycans. CONCLUSIONS – These results suggest that synthetic glycans could be a promising novel anticancer agent for performing chemotherapy against glioblastoma.”
Epidemiological studies
The bottom line is that glucosamine supplementation lowers human mortality
Glucosamine and Chondroitin Sulfate have already been shown in a large epidemiological study to lower overall mortality and reduce cancer risks – a 5-year study of 77,719 elderly residents of Washington State. We found several publications based on different analyses of data based on this population.
The 2010 publication Total mortality risk in relation to use of less-common dietary supplementsreported:“Background: Dietary supplement use is common in older US adults; however, data on health risks and benefits are lacking for a number of supplements. Objective: We evaluated whether 10-y average intakes of 13 vitamin and mineral supplements and glucosamine, chondroitin, saw palmetto, Ginko biloba, garlic, fish-oil, and fiber supplements were associated with total mortality. Design: We conducted a prospective cohort study of Washington State residents aged 50–76 y during 2000–2002. Participants (n= 77,719) were followed for mortality for an average of 5 y. Results:A total of 3577 deaths occurred during 387,801 person-years of follow-up. None of the vitamin or mineral 10-y average intakes were associated with total mortality. Among the nonvitamin-nonmineral supplements, only glucosamine and chondroitin were associated with total mortality. The hazard ratio (HR) when persons with a high intake of supplements (≥4 d/wk for ≥3 y) were compared with nonusers was 0.83 (95% CI: 0.72, 0.97;Pfor trend = 0.009) for glucosamine and 0.83 (95% CI: 0.69, 1.00;Pfor trend = 0.011) for chondroitin. There was also a suggestion of a decreased risk of total mortality associated with a high intake of fish-oil supplements (HR: 0.83; 95% CI: 0.70, 1.00), but the test for trend was not statistically significant. Conclusions:For most of the supplements we examined, there was no association with total mortality. Use of glucosamine and use of chondroitin were each associated with decreased mortality.”
The reported hazard ratio for mortality reduction associated with GlcN consumption is reduction: 0.83. This means that the probability of death for those consuming glucosamine per unit time is 83% of that of controls. In my mind this is impressive
The second publication related to the same epidemiological study is from 2012: Use of glucosamine and chondroitin in relation to mortality: “Glucosamine and chondroitin are products commonly used by older adults in the US and Europe. There is limited evidence that they have anti-inflammatory properties, which could provide risk reduction of several diseases. However, data on their long-term health effects is lacking. To evaluate whether use of glucosamine and chondroitin are associated with cause-specific and total mortality. Participants (n = 77,510) were members of a cohort study of Washington State (US) residents aged 50–76 years who entered the cohort in 2000–2002 by completing a baseline questionnaire that included questions on glucosamine and chondroitin use. Participants were followed for mortality through 2008 (n = 5,362 deaths). Hazard ratios (HR) for death adjusted for multiple covariates were estimated using Cox models. Current (baseline) glucosamine and chondroitin use were associated with a decreased risk of total mortality compared to never use. The adjusted HR associated with current use of glucosamine (with or without chondroitin) was 0.82 (95 % CI 0.75–0.90) and 0.86 (95 % CI 0.78–0.96) for chondroitin (included in two-thirds of glucosamine supplements). Current use of glucosamine was associated with a significant decreased risk of death from cancer (HR 0.87 95% CI 0.76–0.98) and with a large risk reduction for death from respiratory diseases (HR 0.59 95% CI 0.41–0.83). Use of glucosamine with or without chondroitin was associated with reduced total mortality and with reductions of several broad causes of death. Although bias cannot be ruled out, these results suggest that glucosamine may provide some mortality benefit.”
This time the hazard ratio for mortality reduction is .82: 18% fewer deaths per unit of time for those consuming the supplement.
Both of these papers were done on the same group of 77,719 people. We note that this is as great an effect as combining vegan diet and fish consumption! We find this quite surprising. Glucosamine appears to have a comparable or greater effect on mortality reduction and lifespan extension than Meformin, Rapamycin, 2DG, Veganism, and Resveratrol in nematodes and rodents.
Going further based on the same population, a 2013 publication looked at the epidemiological consequences of glucosamine consumption on risk of incurring correctal cancer. Use of glucosamine and chondroitin supplements and risk of colorectal cancer. “PURPOSE: Glucosamine and chondroitin are non-vitamin, non-mineral supplements which have anti-inflammatory properties. These supplements are typically used for joint pain and osteoarthritis and are commonly taken as either glucosamine alone or glucosamine plus chondroitin. An exploratory analysis conducted within the VITamins And Lifestyle (VITAL) study observed any use of glucosamine and chondroitin to be associated with reduced risk of colorectal cancer (CRC) after 5 years of follow-up. METHODS: With two additional years of follow-up, we have studied these associations in greater depth, including associations by frequency/duration of use and by formulation, and have evaluated whether observed associations are modified by factors associated with inflammation. Participants include 75,137 western Washington residents aged 50-76 who completed the mailed VITAL questionnaire between 2000 and 2002. Use of glucosamine and chondroitin was ascertained by questions about supplement use during the 10-year period prior to baseline, and participants were followed for CRC through 2008 (n = 557). Cox regression was used to estimate hazard ratios (HRs) and 95 % confidence intervals (CIs). RESULTS: Persons reporting use of glucosamine + chondroitin on 4+ days/week for 3+ years had a non-statistically significant 45 % lower CRC risk than non-users (HR: 0.55; 95 % CI 0.30-1.01; p-trend: 0.16). This association varied by body mass index (p-interaction: 0.006), with inverse association observed among the overweight/obese (p-trend: 0.02), but not among the underweight/normal weight. Use of glucosamine alone was not significantly associated with CRC risk. CONCLUSIONS: There is great need to identify safe and effective cancer preventive strategies, suggesting that glucosamine and chondroitin may merit further attention as a potential chemopreventive agent.”
Again, we are impressed by the 45% risk reduction for CRC.
Epidemiological evidence exists that glucosamine and chondroitin supplementation reduces inflammatory biomarkers.
The recent April 2014 publication Associations Between Glucosamine and Chondroitin Supplement Use and Biomarkers of Systemic Inflammation looks at the same population from a different viewpoint and reports: “Objectives: Glucosamine and chondroitin supplements have been shown to have anti-inflammatory properties in both in vitro studies and animal models; however, little is known about these relationships in humans. The VITamins and Lifestyle (VITAL) biomarker study evaluated the associations between use of these supplements and a panel of circulating inflammatory biomarkers. Design: Study participants included 217 men and women age 50-75 years living in the Seattle metropolitan area. Use of glucosamine and chondroitin supplements was ascertained by home interview/supplement inventory. Inflammation was assessed by using blood and urine collected at the time of home interview. Measures of systemic inflammation included plasma high-sensitivity C-reactive protein (hsCRP), interleukin (IL)-1β, IL-6, IL-8, tumor necrosis factor (TNF)-α, soluble TNF receptors I and II, and urinary prostaglandin E2-metabolite (PGE-M). Multivariate-adjusted linear regression was used to evaluate the associations between supplement use and biomarkers of inflammation. Results: High users (14 or more pills/week) of chondroitin had 36% lower hsCRP (ratio, 0.64; 95% confidence interval [CI], 0.39-1.04; p for trend=.03) and 27% lower PGE-M (ratio, 0.73; 95% CI, 0.5-0.98; p for trend=.07) than nonusers. Compared with nonusers, high users of glucosamine had 28% lower hsCRP (ratio, 0.72; 95% CI, 0.47-1.08; p for trend=.09) and 24% lower PGE-M (ratio, 0.76; 95% CI, 0.59-0.97; p for trend=0.10). Use of glucosamineand chondroitin supplements was not associated with the other markers of inflammation. Conclusions: These results support prior research suggesting that use of glucosamine and chondroitin is associated with reduced hsCRP and PGE2, but further work is needed to more definitively evaluate the anti-inflammatory potential of these supplements.”
Glucosamine supplementation may be protective against lung cancer.
Data from the same Washington State cohort also provided the basis for the 2011 publication Use of glucosamineand chondroitin and lung cancer risk in the VITamins And Lifestyle (VITAL) cohort. “OBJECTIVE: Inflammation plays an important role in lung carcinogenesis. Epidemiologic studies have reported inverse associations of non-steroidal anti-inflammatory drug (NSAID) use and lungcancerrisk. Previously, we found that ever use ofglucosamineand chondroitin, which have anti-inflammatory properties, were inversely associated with lungcancerrisk. After an additional year of follow-up, we further examined the association including frequency/duration of use, interaction with factors associated with inflammation, and lungcancerhistology. METHODS: Participants were members of the VITamins And Lifestyle cohort. Adults, aged 50-76 years, who were residents of western Washington State, completed a baseline questionnaire in 2000-2002 (n = 76,904). Participants were queried on their use of glucosamine and chondroitin, over the 10 years prior to baseline, and categorized as nonuser, low use < 4 days/week or < 3 years, or high use ≥ 4 days/week and ≥ 3 years. Lung ases (n = 808) were ascertained through linkage to the Surveillance, Epidemiology, and End Resultscancer registry. RESULTS: High 10-year use ofglucosamine[hazard ratio (HR), 0.77; 95% CI: 0.56-1.07; p trend = 0.04] but not chondroitin was associated with a reduction in lungcancer risk. The association with glucosamine was limited to adenocarcinoma (HR, 0.49; 95% CI: 0.27-0.90; p trend <0.01) and was not modified by NSAID use or smoking status. CONCLUSIONS: Our results for glucosamineuse are similar to the prior human studies of NSAID use and lungcancer, both in magnitude and the limitation of the association to adenocarcinoma. Unlike NSAIDs, glucosaminehas no known adverse effects. Although confirmatory studies are needed, glucosamine is an attractive candidate for lung cancer chemoprevention.
It is interesting to note that in all of these studies related to the beneficial effects of glucosamine on mortality and cancers, the VITAL cohort consists of older people between 50 and 76.
The effectiveness of glucosamine for treating osteoarthritis remains a matter of controversy.
While the evidence for glucosamine being life-extending and cancer preventing, earlier studies relating to its effectiveness in treating osteoarthritis have been challenged. The bottom-line on the effectiveness of this approach – the main reason why millions have been taking the supplement – appears to be not really in. The 2014 publication Glucosamine for Osteoarthritis: Biological Effects, Clinical Efficacy, and Safety on Glucose Metabolismreports: “Osteoarthritis is a chronic degenerative disorder that currently represents one of the main causes of disability within the elderly population and an important presenting complaint overall. The pathophysiologic basis of osteoarthritis entails a complex group of interactions among biochemical and mechanical factors that have been better characterized in light of a recent spike in research on the subject. This has led to an ongoing search for ideal therapeutic management schemes for these patients, where glucosamine is one of the most frequently used alternatives worldwide due to their chondroprotective properties and their long-term effects. Its use in the treatment of osteoarthritis is well established; yet despite being considered effective by many research groups, controversy surrounds their true effectiveness. This situation stems from several methodological aspects which hinder appropriate data analysis and comparison in this context, particularly regarding objectives and target variables. Similar difficulties surround the assessment of the potential ability of glucosamine formulations to alter glucose metabolism. Nevertheless, evidence supporting diabetogenesis by glucosamine remains scarce in humans, and to date, this association should be considered only a theoretical possibility.”So, we may have a situation where millions of people who are taking glucosamine is a supplement are probablysignificantly benefiting from it, but they may be taking it for the wrong reason
There may be a downside to taking too much glucosamine
A 2010 publication suggests that taking high doses of glucosamine or prolonged use it glucosamine reduces levels of SIRT1, leads to apoptosis of pancreatic cells and could increase the risk of developing diabetes. Hexosamines stimulate apoptosis by altering T1 action and levels in rodent pancreatic β-cells: “The activity and levels of SIRT1, which promotes cell survival in several models, are linked to glucose concentrations and cellular energy metabolism. The present study aimed to determine whether impaired Sirt1 activity is involved in the induction of apoptosis by the nutrient-sensing hexosamine biosynthesis pathway (HBP). Pancreatic Nit-1, Rin-m5F, and Min6 β-cells were acutely treated at different doses and times with glucosamine, which enters and stimulates the HBP. Sirt1 levels were genetically modulated by retroviral infection. Expression levels, cellular localization, and activity of apoptosis-related markers were determined by qPCR, immunoblotting, and co-immunoprecipitation. Glucosamine treatment dose- and time dependently induced cell apoptosis in all cell lines studied. HBP stimulation time dependently modified SIRT1 protein levels, notably in the cytoplasm. This was concomitant with increased E2F1 binding to the c-myc promoter. In both NIT-1 and min6 β-cells, genetic knockdown of Sirt1 expression resulted in higher susceptibility to HBP-stimulated apoptosis, whereas overexpression of Sirt1 had the opposite impact. These findings indicate that reduction of SIRT1 levels by hexosamines contributes to β-cell apoptosis. Methods to increase SIRT1 levels or activity could thus prevent the decrease in β-cell mass, notably that observed in type 2 diabetes.”
Relative to this issue, the 2014 publication 17A-Estradiol protects against glucosamine-induced pancreatic A-cell dysfunction concludes “: GlcN impairs insulin secretion of [beta]-cells by inhibiting Ca2+ influx and enhancing [beta]-cell apoptosis with increases in ER stress-related proteins, whereas E2 counters these adverse effects of GlcN.”
Glucosamine supplementation and other research approaches to mitochondrial health
There has been a wealth of research going on recently related to mitochondrial health and interventions that could enhance it. To put the above results related to glucosamine in context, I comment very briefly here on some of the “traditional” and “newer” health supplements and interventions intended to impact on the mitochondria.
Glucosamine – activates AMPK, and NRF2, enhances amino acid transport, inhibits glycolysis, mitigates against Warburg, activates autophagy – as above.
Co-enzyme Q10 (ubiquinone)- A powerful mitochondrial endogenous antioxidant. Extremely popular as a dietary supplement, often prescribed, consumed by millions, a $700 million worldwide business. Thought to reduce excess mitochondrial ROS. But currently there is much skepticism as to effectiveness of exogenous administration because of very limited bioavailability (ref)(ref)
Melatonin – This is a circadian-activated hormone that exercises multiple regulatory functions over metabolism. Found with high concentration in mitochondria, it is known primarily for its antioxidant properties. Thought to increase OXPHOS efficiency, reduce mitochondrial membrane potential and mildly promote glycolysis, it transiently raises not only the expression of neuronal NOS mRNA, nNOS protein synthesis and the nitric oxide oxidation products, nitrite and nitrate(ref)(ref).
Acetyl-l-carnitine and alpha-lipoic acid. The Juvenon combination of bioavailable antioxidants is purported to cross the blood-brain barrier and penetrate the mitochondrial membranes. Presumably, they function as antioxidants to reduce excessive ROS produced by old and inefficient mitochondria, and reduce excess mitochondrial membrane polarization(ref).
PQQ – This is a redox cofactor that activates PGC1alpha and the mitochondrial sirtuin SIRT3. Above, we mentioned how PGC1alpha is essential for mitochondrial biogenesis(ref).
Exercise – activates AMPK, PGC1alpha, mitochondrial biogenesis, and autophagy(ref)(ref).
Calorie restriction – Calorie restriction in this various forms, including intermittent fasting, upgrades expression of AMPK with its downstream benefits on mitochondria such as those described above including mitochondrial biogenesis(ref)(ref). Both exercise and diet regulate expression of sirtuins, particularly SIRT1 and SIRT3. SIRT3 is a mitochondrial protein important for deacelating acetyllysine-modified proteins in mitochondria.
NMN (Nicotinamide Mononucleotide) - Assuring adequate levels of nuclear NAD+ is essential for the prevention of a state of pseudo-hypoxia in the nucleus with the result being an adequate generation of mitochondrial proteins essential for averting cellular relapse into Warburg–type metabolism and a number of associated age-related problems such as hearing loss(ref). DNA repair machinery (PARPs) also draws down on NAD+ and it is therefore in short supply with aging. NAD+ is a cofactor for both Sirtuins(and PARPs). There are 7 Sirtuins and over 30 different PARPs. NAD+ is an “on switch” for part of the mitochondrial encoded genes required for Complex II and Complex IV in the mitochondrial electron transfer chain. One thought is that supplementation with NMN could enhance production of NAD+ and be very useful both for DNA repair and mitochondrial health, But this is an extremely expensive substance at the present time(ref)(ref). Our blog was one of the first to suggest NMN supplementation in the 2012 entry by Victor Circadian Regulation,NMN, Preventing Diabetes, and Longevity There is a great deal of current research in this area.
Image sourceImage source NMN promotes nuclear NAD+ which upregulates expression of SIRT1 averting the Warburg effect and Type 2 diabetes.
NR (nicotinamide riboside) - This is a readily available supplement that is also sought possibly to be an effective promoter of nuclear NAD+. Actually. NR is the intermediary in the NMN inducement of NAD+ (ref)(ref)
Resveratrol – promotes expression of SIRT1 needed for mitochondrial health and the mitochondrial proteins SIRT3-4-5. Further, it is an activator of AMPK and autophagy(ref)(ref).
Centrophenoxine (meclophenoxate) – this is an older drug still used in Europe to treat forms of dementia. About half the research publications on it were published prior to 1980(ref) and my impression is that few researchers have seriously looked at it from a contemporary understanding of mitochondria and molecular biology. So, while it is known to have major impacts on cell metabolism, exactly how it works with respect to mitochondria is unclear to me. Among its impacts (disputed) is clearing of lipofuscin from cells(ref)(ref)(ref). This might have a positive impact on mitophagy.
C60 fullerenes in olive oil – functions as antioxidant to reduce excessive ROS produced by old and inefficient mitochondria, reduces excess mitochondrial membrane polarization(ref)(ref).
MitoQ – Co-enzyme Q-10 molecularly modified by adding a charged anion designed to facilitate it penetrating the mitochondrial membrane. Functions as antioxidant to reduce excessive ROS produced by old and inefficient mitochondria, reduces excess membrane polarization(ref)(ref).
SkQ -This list is not exhaustive; other supplements have been researched such as SkQ, like MitoQ, a charged antioxidant molecule designed so it can effectively traverse the mitochondrial membrane(ref)
We seriously question whether certain of these approaches are compatible. Specifically the demonstrated results with glucosamine suggest that ROS plays a very important signaling function in achieving health and longevity benefits via mitohormesis. Therefore, the exogenously administered antioxidants could interfere with such signaling and produce negative impacts for normal healthy people. On the other hand, strong mitochondrial antioxidants could be useful for people whose mitochondria are old and leaky and are already heavily spinning out ROS.
The purpose of mentioning these alternative approaches to mitochondrial health is not to compare their efficacy or characterize what they really do. It is to establish a single point: their mechanisms of operation appear to be very different. Therefore we wonder about which approaches are effective under what conditions, which approaches are compatible, and whether adopting multiple approaches to mitochondrial health might produce benefits that are additive or even possibly multiplicative for healthy people. We intend to explore the rich possibilities inherent in this question in subsequent blog entries.
Trehalose is a natural disaccharide sugar contained in mushrooms and many other organisms. It has remarkable health-producing and life-extending properties that work in mice and lower organisms. That is why we choose to focus on it here. This blog entry details animal model research which suggests significant health and longevity advantages of trehalose consumption. We describe issues related to its bioavailability and safety in humans and wonder why it is not in clinical trials for neurodegenerative diseases.
Basics About trehalose:
“In nature, Trehaose can be found in animals, plants, and microorganisms. In animals, trehalose is prevalent in shrimp, and also in insects, including grasshoppers, locusts, butterflies, and bees, in which blood-sugar is trehalose. The trehalose is then broken down into glucose by the catabolic enzyme trehalase for use. Trehalose is also present in the nutrition exchange liquid of hornets and their larvae(ref).” This is unlike the situation in us humans where our primary blood sugar is glucose. When we eat trehalose, some or much of it is broken down into glucose by the trehalase enzyme in the gut before it enters our bloodstreams.
The enzyme trehalase converts trehalose to glucose in humans and most animals
“In general, the fate of ingested or parenterally administered trehalose corresponds to that of glucose since trehalose is rapidly hydrolysed to glucose by the enzyme trehalase. Trehalase is found in humans and most animals at the brush border of the intestinal mucosa, as well as in the kidney, liver, and blood plasma (Hore & Messer, 1968; van Handel, 1970; Demelier et al., 1975; Labat-Robert, 1982; Niederst & Dauça, 1985; Eze, 1989; Riby et al., 1990; Yoshida, 1993). Trehalase activity has been found in the small intestine of humans, mice, rats, guinea-pigs, rabbits, pigs, and baboons (Cerda et al., 1972; Hietanen, 1973; Ruppin et al., 1974; Maestracci, 1976; Garland, 1989)(from WHO FOOD ADDITIVES SERIES 46:TREHALOSE). The implication is that conversion of trehalose to glucose starts in the gut and normally is continued and finished elsewhere. I am not clear how much of trehalose is converted where under what circumstances. In animal experiments to be described, ingested trehalase has impacts on cells quite opposite of those of glucose impacts.
Trehalose is safe and non-toxic for humans – when eaten
It has been used as a food sweetener in Japan for many years. A January article in Food Chemistry showed that in mice, doses of 1.25gms/kg, 2.5gms/kg, and 5gms/kg were safe Multiple toxicity studies of trehalose in mice by intragastric administration: “The results obtained in the present study could support the conclusion that consumption of trehalose has no adverse effects for humans.” This article suggests that as much as 5gms per day is safe in humans: Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies reports: “No consistent treatment-related, dose-dependent adverse effects were observed in any of the eight safety studies performed at doses up to 10% of the diets. On the basis of these toxicity studies, human studies in which doses of trehalose were administered to various populations, and consumption of trehalose in commercial products in Japan, it is concluded that trehalose is safe for use as an ingredient in consumer products when used in accordance with current Good Manufacturing Practices.”
We note that non-toxicity is probably a corollary of well-functioning of the trehalse enzyme. Evolution invented trehalase in the gut to convert trehalose into glucose before it enters the blood stream. So, administering trehalose intravenously or via liposomes might or might not be so safe. We discuss this issue further below.
Mushrooms are a great source of Trehalose
They sometimes call trehalose “mushroom sugar.” Here are the main edible mushrooms that have trehalose
They can contain 1% to 17% percent of trehalose in dry weight form.
Trehalose stabilizes protein structures
For a long time, trehalose was seen mainly as yet-another sugar occurring in nature. Now. it is known to play very important roles in stabilizing protein structures, the root reason why it is of interest for health and longevity.
The 2009 publication Effect of trehalose on protein structure tells this story: “Trehalose is a ubiquitous molecule that occurs in lower and higher life forms but not in mammals. Till about 40 years ago, trehalose was visualized as a storage molecule, aiding the release of glucose for carrying out cellular functions. This perception has now changed dramatically. The role of trehalose has expanded, and this molecule has now been implicated in a variety of situations. Trehalose is synthesized as a stress-responsive factor when cells are exposed to environmental stresses like heat, cold, oxidation, desiccation, and so forth. When unicellular organisms are exposed to stress, they adapt by synthesizing huge amounts of trehalose, which helps them in retaining cellular integrity. This is thought to occur by prevention of denaturation of proteins by trehalose, which would otherwise degrade under stress. This explanation may be rational, since recently, trehalose has been shown to slow down the rate of polyglutamine-mediated protein aggregation and the resultant pathogenesis by stabilizing an aggregation-prone model protein. In recent years, trehalose has also proved useful in the cryopreservation of sperm and stem cells and in the development of a highly reliable organ preservation solution. This review aims to highlight the changing perception of the role of trehalose over the last 10 years and to propose common mechanisms that may be involved in all the myriad ways in which trehalose stabilizes protein structures. These will take into account the structure of trehalose molecule and its interactions with its environment, and the explanations will focus on the role of trehalose in preventing protein denaturation.” There is much continuing research on trehalose since 2009. Pubmed.org lists 10,428 research publication citations for trehalose
Image may be NSFW. Clik here to view.Hayashibara and Cargill have made a food sweetener product out of Trehalose called TREHA, and they received approval last year for it to be marketed in Canada. It has a relative sweetness of 40-45%, compared to sucrose. Hayashibara makes this in vats from food-grade starch. A one-pound jar of “pure trehalose” can be purchased online for $9.99(ref).
Research findings make trehalose very interesting
A substantial portion of the research literature on trehalose relates primarily to plants, chemical relationships, food preservation and industrial processes. We focus here on items related to health.
1. Trehalose enhances longevity by inhibiting the Insulin/IGF axis in nematodes.
The IGF1 pathway activated by insulin is know to be associated with accelerated aging and age-related degenerative diseases, and is frequently activated in older people, though it plays important roles in all stages of human development. In fact we believe that activation of this pathway is a root cause of aging. The 2010 publication Trehalose extends longevity in the nematode Caenorhabditis elegans reports: “Trehalose is a disaccharide of glucose found in diverse organisms and is suggested to act as a stress protectant against heat, cold, desiccation, anoxia, and oxidation. Here, we demonstrate that treatment of Caenorhabditis elegans with trehalose starting from the young-adult stage extended the mean life span by over 30% without any side effects. Surprisingly, trehalose treatment starting even from the old-adult stage shortly thereafter retarded the age-associated decline in survivorship and extended the remaining life span by 60%. Demographic analyses of age-specific mortality rates revealed that trehalose extended the life span by lowering age-independent vulnerability. Moreover, trehalose increased the reproductive span and retarded the age-associated decrease in pharyngeal-pumping rate and the accumulation of lipofuscin autofluorescence. Trehalose also enhanced thermotolerance and reduced polyglutamine aggregation. These results suggest that trehalose suppressed aging by counteracting internal or external stresses that disrupt protein homeostasis. On the other hand, the life span-extending effect of trehalose was abolished in long-lived insulin/IGF-1-like receptor (daf-2) mutants. RNA interference-mediated inactivation of the trehalose-biosynthesis genes trehalose-6-phosphate synthase-1 (tps-1) and tps-2, which are known to be up-regulated in daf-2 mutants, decreased the daf-2 life span. These findings indicate that a reduction in insulin/IGF-1-like signaling extends life span, at least in part, through the aging-suppressor function of trehalose. Trehalose may be a lead compound for potential nutraceutical intervention of the aging process.”
Although other interventions in nematodes can do more, 60% life extension in older animals is not too shabby and led us to wonder further about exactly how trehalose works.
2. Trehalose induces autophagy independently of mTOR pathway inhibition and has been shown in experimental models to help in neurodegenerative diseases. Trehalose can clear alpha-synuclein, polyglutamate, amyloid beta 40, amyloid beta 42, and tau proteins such as this seen in Huntington’s disease, Parkinson’s disease, and Alzheimer’s disease.
A series of publications going back to 2004 establish these important points.
The 2004 publication Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease suggested that the beneficial actions of trehalose in neurodegenerative diseases could be as a chaperone molecule affecting protein folding. It reported: “Inhibition of polyglutamine-induced protein aggregation could provide treatment options for polyglutamine diseases such as Huntington disease. Here we showed through in vitro screening studies that various disaccharides can inhibit polyglutamine-mediated protein aggregation. We also found that various disaccharides reduced polyglutamine aggregates and increased survival in a cellular model of Huntington disease. Oral administration of trehalose, the most effective of these disaccharides, decreased polyglutamine aggregates in cerebrum and liver, improved motor dysfunction and extended lifespan in a transgenic mouse model of Huntington disease. We suggest that these beneficial effects are the result of trehalose binding to expanded polyglutamines and stabilizing the partially unfolded polyglutamine-containing protein. Lack of toxicity and high solubility, coupled with efficacy upon oral administration, make trehalose promising as a therapeutic drug or lead compound for the treatment of polyglutamine diseases. The saccharide-polyglutamine interaction identified here thus provides a new therapeutic strategy for polyglutamine diseases.”
In the pancreas, insulin forms a type of amyloid called “amylin.” It is essentially misfolded insulin molecule. Trehalose prevents this from occurring. In this case it may be functioning as a “chemical chaperone,” The 2004 publication Inhibition of insulin amyloid formation by small stress moleculesreported: “Amyloidogenic proteins undergo an alternative folding pathway under stressful conditions leading to formation of fibrils having cross beta-sheet structure, which is the hallmark of many neurodegenerative diseases. As a means of surviving against external stress, on the other hand, many microorganisms accumulate small stress molecules to prevent abnormal protein folding and to contribute to protein stability, which hints at the efficacy of the solutes against amyloid formation. The current work demonstrates the effectiveness of small stress molecules such as ectoine, betaine, trehalose, and citrulline on inhibition of insulin amyloid formation in vitro. The inhibitory effects were analyzed by thioflavin T-induced fluorescence, circular dichroism, and atomic force microscopy. This report suggests that naturally occurring small molecules may serve a function that is typically fulfilled by protein chaperones, and it provides a hint for designing inhibitors against amyloid formation associated with neurodegenerative disorders.
A 2005 publication Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42 relates to a possible role for trehalose in preventing or treating Alzheimer’s disease. “These results also suggest that the use of trehalose, a highly soluble, low-priced sugar, as part of a potential therapeutic cocktail to control Abeta peptide aggregation and toxicity warrants further investigation.”
The 2006 publication Trehalose, a Novel mTOR-independent Autophagy Enhancer, Accelerates the Clearance of Mutant Huntingtin and α-Synuclein points out how its autophagic effects are mTOR-independent. “Trehalose, a disaccharide present in many non-mammalian species, protects cells against various environmental stresses. Whereas some of the protective effects may be explained by its chemical chaperone properties, its actions are largely unknown. Here we report a novel function of trehalose as an mTOR-independent autophagy activator. Trehalose-induced autophagy enhanced the clearance of autophagy substrates like mutant huntingtin and the A30P and A53T mutants of α-synuclein, associated with Huntington disease (HD) and Parkinson disease (PD), respectively. Furthermore, trehalose and mTOR inhibition by rapamycin together exerted an additive effect on the clearance of these aggregate-prone proteins because of increased autophagic activity. By inducing autophagy, we showed that trehalose also protects cells against subsequent pro-apoptotic insults via the mitochondrial pathway. The dual protective properties of trehalose (as an inducer of autophagy and chemical chaperone) and the combinatorial strategy with rapamycin may be relevant to the treatment of HD and related diseases, where the mutant proteins are autophagy substrates.”
The 2010 publication Trehalose ameliorates dopaminergic and tau pathology in parkin deleted/tau overexpressing mice through autophagy activation reports: “Tauopathies are neurodegenerative diseases, sporadic or familial, mainly characterized by dementia and parkinsonism associated to atrophy of the frontotemporal cortex and the basal ganglia, with deposition of abnormal tau in brain. Hereditary tauopathies are related with mutations of the tau gene. Up to the present, these diseases have not been helped by any disease-modifying treatment, and patients die a few years after the onset of symptoms. — We have developed and characterized a mouse model of tauopathy with parkinsonism, overexpressing human mutated tau protein with deletion of parkin (PK−/−/TauVLW). At 3 months of age, these mice present abnormal dopamine-related behavior, severe dropout of dopamine neurons in the ventral midbrain, reduced dopamine levels in the striatum and abundant phosphorylated tau-positive neuritic plaques, neurofibrillary tangles, astrogliosis, and, at 12 months old, plaques of murine β-amyloid in the hippocampus. — Trehalose is a natural disaccharide that increases the removal of abnormal proteins through enhancement of autophagy. In this work, we tested if 1% trehalose in the drinking water reverts the PK−/−/TauVLWphenotype. The treatment with trehalose of 3-month-old PK−/−/TauVLW mice for 2.5 months reverted the dropout of dopamine neurons, which takes place in the ventral midbrain of vehicle treated PK−/−/TauVLWand the reduced dopamine-related proteins levels in the midbrain and striatum. The number of phosphorylated tau-positive neuritic plaques and the levels of phosphorylated tau decreased, as well as astrogliosis in brain regions. The autophagy markers in the brain, the autophagic vacuoles isolated from the liver, and the electron microscopy data indicate that these effects of trehalose are mediated by autophagy. The treatment with trehalose for 4 months of 3-month-old PK−/−/TauVLW mice maintained the amelioration of the tau pathology and astrogliosis but failed to revert DA-related pathology in the striatum. Furthermore, the 3-week treatment with trehalose of 14-month-old PK−/−/TauVLW mice, at the limit of their life expectancy, improved the motor behavior and anxiety of these animals, and reduced their levels of phosphorylated tau and the number of murine β-amyloid plaques. — Trehalose is neuroprotective in this model of tauopathy. Since trehalose is free of toxic effects at high concentrations, this study opens the way for clinical studies of the effects of trehalose in human tauopathies.”
The 2010 publication Chemical Inducers of Autophagy That Enhance the Clearance of Mutant Proteins in Neurodegenerative Diseases instead suggests that the action of trehalose is via autophagy. It has to say: “Another chemical screen to identify inhibitors of polyQ-mediated protein aggregation in vitro identified t.rehalose (a disaccharide) as an inhibitor of mutant huntingtin aggregation, which reduced toxicity in HD cell models and attenuated disease pathology in a mouse model of HD (59). This protective effect of trehalose was suggested to be mediated by its ability to act as a chemical chaperone through its binding to the polyQ-expanded mutant huntingtin and influencing its protein folding and aggregation. However, we have shown that trehalose enhances the autophagy pathway independently of mTOR, thereby increasing the clearance of mutant aggregate-prone proteins (60). Additionally, trehalose protected against pro-apoptotic insults via autophagy (60). The myriad of protective properties of trehalose acting as an autophagy inducer and chemical chaperone, coupled with its lack of toxicity, may be of benefit in the treatment of neurodegenerative disorders.” The critical importance of autophagy is detailed in our 2013 blog entry Autophagy – the housekeeper in every cell that fights aging.
The 2011 article The accumulation of neurotoxic proteins, induced by proteasome inhibition, is reverted by trehalose, an enhancer of autophagy, in human neuroblastoma cells reports: “Neurodegenerative diseases like Parkinson’s disease, Alzheimer’s disease, Huntington’s disease and others are due to accumulation of abnormal proteins which fold improperly and impair neuronal function. Accumulation of these proteins could be achieved by several mechanisms including mutation, overproduction or impairment of its degradation. Inhibition of the normal protein degradation is produced by blockade of the ubiquitin proteasome system. — We have shown that epoxomicin, a proteasome inhibitor, increases the levels of proteins involved in neurodegenerative disorders such as α-synuclein and hyper phosphorylated tau in NB69 human neuroblastoma cells and that such increase correlates with an enhanced rate of cell death. We then investigated whether the stimulation of autophagy, an alternative mechanism for elimination of abnormal proteins, by treatment with trehalose, counteracts the effects of proteasomal blockade. Trehalose, a disaccharide present in many non-mammalian species, known to enhance autophagy, protects cells against various environmental stresses. Treatment with trehalose produced a dose and time-dependent increase in the number of autophagosomes and markers of autophagy in NB69 cells. Trehalose did not change the number of total neither the number of dividing cells in the culture but it completely prevented the necrosis of NB69 induced by epoxomicin. In addition, the treatment with trehalose reverted the accumulation, induced by epoxomicin, of polyubiquitinated proteins, total and phosphorylated tau, p-GSK-3, and α-synuclein, as well as the α-synuclein intracellular aggregates. The effects of trehalose were not mediated through activation of free radical scavenging compounds, like GSH, or mitochondrial proteins, like DJ1, but trehalose reduced the activation of ERK and chaperone HSP-70 induced by epoxomicin. Inhibition of ERK phosphorylation prevented the epoxomicin-induced cell death. Inhibition of autophagy reverted the neuroprotective effects of trehalose in epoxomicin-induced cell death. These results suggest that trehalose is a powerful modifier of abnormal protein accumulation in neurodegenerative diseases.” Because this experiment was with human cells, the suggestion is that the results observed in mice might well carry over to humans.
The 2012 publication Autophagic degradation of tau in primary neurons and its enhancement by trehalose relates: “Modulating the tau level may represent a therapeutic target for Alzheimer’s disease (AD), as accumulating evidence shows that Abeta-induced neurodegeneration is mediated by tau. It is therefore important to understand the expression and degradation of tau in neurons. Recently we showed that overexpressed mutant tau and tau aggregates are degraded via the autophagic pathway in an N2a cell model. Here we investigated whether autophagy is involved in the degradation of endogenous tau in cultured primary neurons. We activated this pathway in primary neurons with trehalose, an enhancer of autophagy. This resulted in the reduction of endogenous tau protein. Tau phosphorylation at several sites elevated in AD pathology had little influence on its degradation by autophagy. Furthermore, by using a neuronal cell model of tauopathy, we showed that activation of autophagy suppresses tau aggregation and eliminates cytotoxicity. Notably, apart from activating autophagy, trehalose also inhibits tau aggregation directly. Thus, trehalose may be a good candidate for developing therapeutic strategies for AD and other tauopathies.”
Finally, the 2012 publication Stimulation of autophagy reduces neurodegeneration in a mouse model of human tauopathy reports: “The accumulation of insoluble proteins is a pathological hallmark of several neurodegenerative disorders. Tauopathies are caused by the dysfunction and aggregation of tau protein and an impairment of cellular protein degradation pathways may contribute to their pathogenesis. Thus, a deficiency in autophagy can cause neurodegeneration, while activation of autophagy is protective against some proteinopathies. Little is known about the role of autophagy in animal models of human tauopathy. In the present report, we assessed the effects of autophagy stimulation by trehalose in a transgenic mouse model of tauopathy, the human mutant P301S tau mouse, using biochemical and immunohistochemical analyses. Neuronal survival was evaluated by stereology. Autophagy was activated in the brain, where the number of neurons containing tau inclusions was significantly reduced, as was the amount of insoluble tau protein. This reduction in tau aggregates was associated with improved neuronal survival in the cerebral cortex and the brainstem. We also observed a decrease of p62 protein, suggesting that it may contribute to the removal of tau inclusions. Trehalose failed to activate autophagy in the spinal cord, where it had no impact on the level of sarkosyl-insoluble tau. Accordingly, trehalose had no effect on the motor impairment of human mutant P301S tau transgenic mice. Our findings provide direct evidence in favour of the degradation of tau aggregates by autophagy. Activation of autophagy may be worth investigating in the context of therapies for human tauopathies.”
3. Trehalose has also been shown to be effective for treating prion disease.
When prion infected organisms are given trehalose, they start degrading the prions. The response occurred at concentrations of 100 mM – that is not that high!. The 2009 publication Autophagy induction by trehalose counteracts cellular prion infection reports: “Prion diseases are fatal neurodegenerative and infectious disorders for which no therapeutic or prophylactic regimens exist. In search of cellular mechanisms that play a role in prion diseases and have the potential to interfere with accumulation of intracellular pathological prion protein (PrP(Sc)), we investigated the autophagic pathway and one of its recently published inducers, trehalose. Trehalose, an alpha-linked disaccharide, has been shown to accelerate clearance of mutant huntingtin and alpha-synuclein by activating autophagy, mainly in an mTOR-independent manner. Here, we demonstrate that trehalose can significantly reduce PrP(Sc) in a dose- and time-dependent manner while at the same time it induces autophagy in persistently prion-infected neuronal cells. Inhibition of autophagy, either pharmacologically by known autophagy inhibitors like 3-methyladenine, or genetically by siRNA targeting Atg5, counteracted the anti-prion effect of trehalose. Hence, we provide direct experimental evidence that induction of autophagy mediates enhanced cellular degradation of prions. Similar results were obtained with rapamycin, a known inducer of autophagy, and imatinib, which has been shown to activate autophagosome formation. While induction of autophagy resulted in reduction of PrP(Sc), inhibition of autophagy increased the amounts of cellular PrP(Sc), suggesting that autophagy is involved in the physiological degradation process of cellular PrP(Sc). Preliminary in vivo studies with trehalose in intraperitoneally prion-infected mice did not result in prolongation of incubation times, but demonstrated delayed
4. Trehalose can regulate lipid metabolism (Macrolipophagy) via autophagy
The 2010 publication Autophagy regulates lipid metabolism reports “The intracellular storage and utilization of lipids are critical to maintain cellular energy homeostasis. During nutrient deprivation, cellular lipids stored as triglycerides in lipid droplets are hydrolysed into fatty acids for energy. A second cellular response to starvation is the induction of autophagy, which delivers intracellular proteins and organelles sequestered in double-membrane vesicles (autophagosomes) to lysosomes for degradation and use as an energy source. Lipolysis and autophagy share similarities in regulation and function but are not known to be interrelated. Here we show a previously unknown function for autophagy in regulating intracellular lipid stores (macrolipophagy). Lipid droplets and autophagic components associated during nutrient deprivation, and inhibition of autophagy in cultured hepatocytes and mouse liver increased triglyceride storage in lipid droplets. This study identifies a critical function for autophagy in lipid metabolism that could have important implications for human diseases with lipid over-accumulation such as those that comprise the metabolic syndrome.”
We did not previously know about this relationship between autophagy and liquid metabolism. Insulin turns off lipid metabolism and turns on lipid storage. This is why people get fat when their insulin levels are high. Secondly, the cited article points out that when autophagy is inhibited, lipids (triglycerides) built up in the liver. Thus “fatty liver” is due to high insulin levels and autophagy inhibition, NOT dietary fat intake. Then they went on to do an experiment directly linking autophagy to lipid metabolism They called this “macrolipophagy.” I (Jim) think this is why the Atkins diet (or ketogenic diet) works!
5.Illustrative of the versatility and multiple mechanisms of operation of trehalose is its operation in cases of vasospasm.
The 2012 publication Trehalose treatment suppresses inflammation, oxidative stress, and vasospasm induced by experimental subarachnoid hemorrhage reports: “Background: Subarachnoid hemorrhage (SAH) frequently results in several complications, including cerebral vasospasm, associated with high mortality. Although cerebral vasospasm is a major cause of brain damages after SAH, other factors such as inflammatory responses and oxidative stress also contribute to high mortality after SAH. Trehalose is a non-reducing disaccharide in which two glucose units are linked by α,α-1,1-glycosidic bond, and has been shown to induce tolerance to a variety of stressors in numerous organisms. In the present study, we investigated the effect of trehalose on cerebral vasospasm, inflammatory responses, and oxidative stress induced by blood in vitro and in vivo. Methods: Enzyme immunoassay for eicosanoids, pro-inflammatory cytokines, and endothelin-1, and western blotting analysis for cyclooxygenase-2, inducible nitric oxide synthase, and inhibitor of NF-κB were examined in macrophage-like cells treated with hemolysate. After treatment with hemolysate and hydrogen peroxide, the levels of lipid peroxide and amounts of arachidonic acid release were also analyzed. Three hours after the onset of experimental SAH, 18 Japanese White rabbits received an injection of saline, trehalose, or maltose into the cisterna magna. Angiographic and histological analyses of the basilar arteries were performed. In a separate study, the femoral arteries from 60 rats were exposed to fresh autologous blood. At 1, 3, 5, 7, 10, and 20days after treatment, cryosections prepared from the femoral arteries were histologically analyzed. Results: When cells were treated with hemolysate, trehalose inhibited the production of several inflammatory mediators and degradation of the inhibitor of NF-κB and also suppressed the lipid peroxidation, the reactive oxygen species-induced arachidonic acid release in vitro. In the rabbit model, trehalose produced an inhibitory effect on vasospasm after the onset of experimental SAH, while maltose had only a moderate effect. When the rat femoral arteries exposed to blood were investigated for 20days, histological analysis revealed that trehalose suppressed vasospasm, inflammatory response, and lipid peroxidation. Conclusions: These data suggest that trehalose has suppressive effects on several pathological events after SAH, including vasospasm, inflammatory responses, and lipid peroxidation. Trehalose may be a new therapeutic approach for treatment of complications after SAH.”
6. Trehalose penetrates the blood-brain barrier quite well.
Since it has been shown to stimulate autophagy and clear protein aggregates in the brain with neurodegenerative diseases, for trehalose to be effective it would be very important for it to cross the blood brain barrier. It does. This was shown in an Alzheimer’s conference a few years ago. It is an effective treatment for animal models of Alzhiemer’s disease even after the onset of the disease. It is also effective to prevent Alzheimer’s disease from occurring. See the 2010 publication A diverse portfolio of novel drug discovery eff orts for Alzheimer’s disease: Meeting report from the 11th International Conference on Alzheimer’s Drug Discovery, 27–28 September 2010, Jersey City, NJ, USA“Trehalose, which has good blood–brain barrier penetration, was effective as both a therapeutic agent after disease onset as well as a long-term preventative treatment that improved motor function in the JNPL3 mouse model.”
This is more good news.
7. While trehalose is a sugar, it does does not form Advanced Glycation Endproducts (AGEs)
This again was news for us, “Trehalose is a nonreducing sugar formed from two glucose units joined by a 1-1 alpha bond, giving it the name of α-D-glucopyranosyl-(1→1)-α-D-glucopyranoside. The bonding makes trehalose very resistant to acid hydrolysis, and therefore is stable in solution at high temperatures, even under acidic conditions. The bonding also keeps nonreducing sugars in closed-ring form, such that the aldehyde or ketone end groups do not bind to the lysine or arginine residues of proteins (a process called glycation)(ref).” Tissue glycation has been seen as a major problem of aging for a very long time as pointed out in my treatise in 2007(ref). This is yet more good news.
8. Trehalose may prevent the progression of insulin resistance
The 2010 publication Trehalose prevents adipocyte hypertrophy and mitigates insulin resistance reports: “Trehalose has been shown to evoke lower insulin secretion than glucose in oral saccharide tolerance tests in humans. Given this hypoinsulinemic effect of trehalose, we hypothesized that trehalose suppresses adipocyte hypertrophy by reducing storage of triglyceride and mitigates insulin resistance in mice fed a high-fat diet (HFD). Mice were fed an HFD and given drinking water containing 2.5% saccharide (glucose [Glc], trehalose [Tre], maltose [Mal], high-fructose corn syrup, or fructose [Fru]) ad libitum. After 7 weeks of HFD and saccharide intake, fasting serum insulin levels in the Tre/HFD group were significantly lower than in the Mal/HFD and Glc/HFD groups (P < .05). Furthermore, the Tre/HFD group showed a significantly suppressed elevation of homeostasis model assessment-insulin resistance compared with the Mal/HFD group (P < .05) and showed a trend toward lower homeostasis model assessment-insulin resistance than the Glc/HFD group. After 8 weeks of feeding, mesenteric adipocyte size in the Tre/HFD group showed significantly less hypertrophy than the Glc/HFD, Mal/HFD, high-fructose corn syrup/HFD, or Fru/HFD group. Analysis of gene expression in mesenteric adipocytes showed that no statistically significant difference in the expression of monocyte chemoattractant protein-1 (MCP-1) messenger RNA (mRNA) was observed between the Tre/HFD group and the distilled water/standard diet group, whereas a significant increase in the MCP-1 mRNA expression was observed in the Glc/HFD, Mal/HFD, Fru/HFD, and distilled water/HFD groups. Thus, our data indicate that trehalose prevents adipocyte hypertrophy and mitigates insulin resistance in HFD-fed mice by reducing insulin secretion and down-regulating mRNA expression of MCP-1. These findings further suggest that trehalose is a functional saccharide that mitigates insulin resistance.”
The suggestion is that if we could substitute trehalose for conventional sugar, that might go a long way towards solving the diabetes epidemic.
0. Trehalose treatment reverses autophagy impairment and prevents neural tube defects in diabetic pregnancies – in mice at least
The 2013 publication Trehalose prevents neural tube defects by correcting maternal diabetes-suppressed autophagy and neurogenesis reports: “Preexisting maternal diabetes increases the risk of neural tube defects (NTDs). The mechanism underlying maternal diabetes-induced NTDs is not totally defined, and its prevention remains a challenge. Autophagy, an intracellular process to degrade dysfunction protein and damaged cellular organelles, regulates cell proliferation, differentiation, and apoptosis. Because autophagy impairment causes NTDs reminiscent of those observed in diabetic pregnancies, we hypothesize that maternal diabetes-induced autophagy impairment causes NTD formation by disrupting cellular homeostasis, leading to endoplasmic reticulum (ER) stress and apoptosis, and that restoration of autophagy by trehalose, a natural disaccharide, prevents diabetes-induced NTDs. Embryos from nondiabetic and type 1 diabetic mice fed with or without 2 or 5% trehalose water were used to assess markers of autophagy, ER stress, and neurogenesis, numbers of autophagosomes, gene expression that regulates autophagy, NTD rates, indices of mitochondrial dysfunction, and neuroepithelial cell apoptosis. Maternal diabetes suppressed autophagy by significantly reducing LC3-II expression, autophagosome numbers, and GFP-LC3 punctate foci in neuroepithelial cells and by altering autophagy-related gene expression. Maternal diabetes delayed neurogenesis by blocking Sox1 neural progenitor differentiation. Trehalose treatment reversed autophagy impairment and prevented NTDs in diabetic pregnancies. Trehalose resolved homeostatic imbalance by correcting mitochondrial defects, dysfunctional proteins, ER stress, apoptosis, and delayed neurogenesis in the neural tubes exposed to hyperglycemia. Our study demonstrates for the first time that maternal diabetes suppresses autophagy in neuroepithelial cells of the developing neural tube, leading to NTD formation, and provides evidence for the potential efficacy of trehalose as an intervention against hyperglycemia-induced NTDs.”
10. Trehalose exercises an antidepressant affect, at least on mice
The 2013 publication Trehalose induced antidepressant-like effects and autophagy enhancement in mice reports: “RATIONALE: The disaccharide trehalose protects cells from hypoxic and anoxic injury and suppresses protein aggregation. In vivo studies with trehalose show cellular and behavioral beneficial effects in animal models of neurodegenerative diseases. Moreover, trehalose was shown to enhance autophagy, a process that had been recently suggested to be involved in the therapeutic action of antidepressant and mood-stabilizing drugs. OBJECTIVE: The present study was therefore designed to explore antidepressant and mood-stabilizing activity of trehalose in animal models for depression and mania. METHODS: Trehalose 1 or 2% was administered for 3 weeks as a drinking solution to Black Swiss mice (a model of manic-like behaviors) or 2% to ICR mice and their behavior evaluated in a number of tests related to depression or mania. The effects of trehalose were compared with similar chronic administration of the disaccharide maltose as well as with a vehicle (water) control. RESULTS: Chronic administration of trehalose resulted in a reduction of frontal cortex p62/beclin-1 ratio suggesting enhancement of autophagy. Trehalose had no mood-stabilizing effects on manic-like behavior in Black Swiss mice but instead augmented amphetamine-induced hyperactivity, an effect similar to antidepressant drugs. In ICR mice, trehalose did not alter spontaneous activity or amphetamine-induced hyperactivity but in two separate experiments had a significant effect to reduce immobility in the forced swim test, a standard screening test for antidepressant-like effects. CONCLUSIONS: The results suggest that trehalose may have antidepressant-like properties. It is hypothesized that these behavioral changes could be related to trehalose effects to enhance autophagy.”
11. There is an issue of how much trehalose ingested by humans makes it into the bloodstream where it can induce the beneficial effects outlined above.
We mentioned that the small intestine has an enzyme called trehalase. This enzyme is also found in the kidney. The trehalase enzyme breaks the trehalose into two separate glucopyranoside sugar molecules that are then absorbed. One gram of trehalose produces 4 kcals/gm of sugar. From Health Canada archived content: “This disaccharide is enzymatically hydrolysed by the enzyme trehalase in the small intestine into two glucose subunits which are subsequently absorbed and metabolized in a manner similar to maltose. Common disaccharides such as sucrose and lactose are digested through the same physiological process. The sugar provides approximately 4 kcal of food energy per gram as do other digestible sugars and starches. It is therefore important that diabetics understand that it is a sugar and for everyone to understand that it contributes Calories. The enzyme trehalase is found in human intestinal system and kidneys although its activity varies and it may be missing in some people. In those cases, malabsorption similar to that found with consumption of lactose in lactose intolerant individuals, or sugar alcohols in most people, may be observed. Fewer persons are thought to be trehalase deficient than are lactase deficient.”
Like humans, mice have trehalase in their guts, developing its expression early in their lives (ref)(ref)(ref). The experiments described above based on adding trehalose to mouse drinking water suggest that significant benefits are nonetheless realized via upgrading autophagy and mitigating insulin resistance(ref)(ref)(ref). So, some trehalose must be getting into cells. I infer this is due to a certain portion of the trehhalose evading gut trehalase breakdown and passing into the bloodstream where it is later broken down by circulating trehalase or in kidneys. I further infer that that the same happens when humans consume trehalose. The percentage amount entering the bloodstream is unknown. However as identified above, it is thought that 5 gms per day is a safe amount for human consumption.
12. Not all sugars are equally healthy. Fructose, in contrast to trehalose, triggers pro-aging and procancer IGF-1 signaling, mTOR activation and inhibition of FOXO3A.
Fructose appears to be a particularly evil sugar. It does this by binding to the GLUT5 transporter. Here is a rather complex diagram on carbohydrate digestion and absorption applicable to humans as well as the naked mole rat.
Image and legend source “Dietary carbohydrate in humans and omnivorous animals is a major nutrient. The carbohydrates that we ingest vary from the lactose in milk to complex carbohydrates. These carbohydrates are digested to monosaccharides, mostly glucose, galactose and fructose, prior to absorption in the small intestine. Glucose and galactose are initially transported into the enterocyte by SGLT1 located in the apical brush border membrane and then exit through the basolateral membrane by either GLUT2 or exocytosis. In a new model of intestinal glucose absorption, transport by SGLT1 induces rapid insertion and activation of GLUT2 in the brush border membrane by a PKC betaII-dependent mechanism. Moreover, trafficking of apical GLUT2 is rapidly up-regulated by glucose and artificial sweeteners, which act through T1R2 + T1R3/alpha-gustducin to activate PLC-beta2 and PKC-beta II. Fructose is transported separately by the brush border GLUT5 and then released out of the enterocyte into the blood by GLUT2.”
Interpretation: fructose is the only sugar absorber by the GLUT5 transporter. Once in the cell fructose directly activates the intracellular portion if the Insulin/IGF-1 pathway (PI3K/PIP3/Akt kinase cascade). The net result of Akt activation is mTOR activation and inhibition of the FOXO transcription factors, of which FOXO3a is a bona fide longevity gene. This is why high fructose corn syrup probably shortens lifespan and decreases health span. It is worse than glucose.
Driving through Iowa a few years back I noticed long train after long train of tank cars and vast sidings loaded with them – all for high fructose corn syrup, not petroleum. Among the hundreds or thousands of processed foods and medicines containing this fructose syrup are the ones on this list.
13. There are issues with using animal knockout models that could lead to lack of translatability of some of the above findings to aging humans
Most of the studies on the beneficial effects of trehalose on protein aggregations in neurodegenerative diseases listed above have been have been based on use of transgenic animal models. That is, on use of probably-younger mice where critical genes have been knocked out so as to lead to an animal model of the human disease to be simulated, be it Huntingtons, Alzheimers, etc. The main beneficial effects seem to be due to induction of autophagy. As far as we can tell, these studies were not specifically on older mice. Maria Cuervo at the Albert Einstein College of Medicine is among those who have written extensively about how aging impedes autophagy by inducing changes in cell lipid membranes. Why chaperone-mediated autophagy tends not to work with aging is because of age-related contamination of liposomal membranes. Therefore, trehalose may not work or work less well to induce autophagy in older animals or people. See Altered dynamics of the lysosomal receptor for chaperone-mediated autophagy with age. “We conclude that age-related changes at the lysosomal membrane are responsible for the altered turnover of the CMA receptor in old organisms and the consequent decline in this pathway.” This age-related effect tcould be due, for example, to consumption of a high fat or cholesterol-rich diet(ref)
On the other hand, there are publications that suggests that age-related decline in autophagy in older mice can be restored via trehalose: The 2012 publication Translational evidence that impaired autophagy contributes to arterial ageing relates: “In old mice, treatment with the autophagy-enhancing agent trehalose restored expression of autophagy markers, rescued NO-mediated EDD by reducing oxidative stress, and normalized inflammatory cytokine expression. In cultured endothelial cells, inhibition of autophagy increased oxidative stress and reduced NO production, whereas trehalose enhanced NO production via an autophagy-dependent mechanism. These results provide the first evidence that autophagy is impaired with ageing in vascular tissues. Our findings also suggest that autophagy preserves arterial endothelial function by reducing oxidative stress and inflammation and increasing NO bioavailability. Autophagy-enhancing therapies may therefore have therapeutic efficacy for ameliorating age-associated arterial dysfunction and preventing CVD.” Also spermadine is among other autophagy-inducers that can reverse arterial aging(ref).
So, it appears to me (Vince) that the bottom line on whether some or all of the beneficial autophshy and IGF-1 suppression effects of trehalose observed in mouse models translate to aged humans is not in.
14. Supplementation with trehalose could lead to people with a rare disease, trehalase deficiency disorder, to have problems.
“Trehalase deficiency is a metabolic condition in which the body is not able to convert trehalose, a disaccharide found in mushrooms, algae and insects, into glucose.[1][2][3] Individuals with this deficiency experience vomiting, abdominal discomfort and diarrhea after eating foods containing trehalose.[4][5]Studies suggest that this condition is very rare in white Americans, very frequent in Greenland natives (where it has been reported in 8% of the population), and not infrequent among Finns.[2][3][4][6]However, since ingestion of large quantities of foods containing trehalose in not common, the real frequency in unknown. Most cases appear to be inherited in an autosomal recessive manner.[2](ref)”
15. Scientific studies clearly establishing health benefits of trehalose supplementation in humans appear to be conspicuously missing
I am frankly puzzled and bothered by the fact that as far as I (Vince) can tell, there are no human studies related to the health and longevity impacts of trehalose other than for the old safety studies and the few cell-level studies mentioned above. So we don’t know whether health benefits can actually be achieved by human consumption of trehalose. One would think that the string of animal studies listed above showing very positive health impacts based on understandable mechanisms of action would lead to a big impetus to see if the same results hold up in humans – and to see whether trehalose can help us with our otherwise intractable neurological and metabolic diseases. And, possibly help is to live longer. But I found no published clinical trials, no population studies and not even any tiny studies testing the consequences of trehelulose consumption by a few people. Note that all the studies I listed above where published in past years. I can understand why drug companies will not invest money in a natural sugar that can’t be patented. But what about university researchers and public health advocates? If it does such great things for mouse health when added to their drinking water, shouldn’t we be thinking of adding it to our water?
I did a search in clinicaltrials,gov using the term “trehalose” and was at first very excited to retrieve 1023 studies in various stages of competion. A closer look, however, revealed that almost all of these retrieved studies are not really about trehalose (e.g. the studies were retrieved because trehalose plays a role as a cancer biomarker) and only 2 or 3 of the studies are actually about testing health benefits of trehalose in humans. One trial in particular, might get the ball rolling. Oral Trehalose Therapy to Reverse Arterial Aging in Middle-Aged and Older Adults, sponsored by the University of Colorado in Boulder. The trial was initiated in 2012 and is still in the recruiting stage. “The proposed study will assess the ability of a naturally occurring sugar to improve the function of arteries with age. Overall, the proposed research project has the long-term potential to influence clinical practice guidelines by establishing a novel, easy to deliver, cost-effective therapy for treating age-associated arterial dysfunction and reducing the risk of cardiovascular disease with age.”
So, is trehalose a sugar mainly interesting because it is absorbed slowly and therefore less likely to initiate sudden glucose level spikes? Or is trehalose a powerful chaperone and autophagy-inducing substance that could be beneficial for the prevention and treatment of major neurological disease in humans? Can consumption of trehalose slow human aging? We don’t know and the way things are going it looks like it it will be a long time before we find out. The research required to answer these questions mostly does not seem to be going on.
Continuing a tradition in this blog of reviewing particularly interesting phytochemicals, we here discuss research on health-related properties and applications of anacardic acid. Although much of the research on anacardic acid has been conducted over a 40 year period, we have not reported on it previously in this blog. Its epigenetic and other properties applicable to medicine are unique and are of high current interest. And there have been important new findings about it in just the last two years.
Basics about anacardic acid
The medicinal use of anacardic acid goes back long before it was called that. “The traditional Ayurveda depicts nutshell oil as a medicinal remedy for alexeritic, amebicidal, gingivitis, malaria and syphilitic ulcers. However, the enduring research and emerging evidence suggests that AA (anacardic acid) could be a potent target molecule with bactericide, fungicide, insecticide, anti-termite and molluscicide properties and as a therapeutic agent in the treatment of the most serious pathophysiological disorders like cancer, oxidative damage, inflammation and obesity. Furthermore, AA was found to be a common inhibitor of several clinically targeted enzymes such as NFκB kinase, histone acetyltransferase (HATs), lipoxygenase (LOX-1), xanthine oxidase, tyrosinase and ureases(ref).”
Modern history of the substance goes back to a publication in 1847 which described an acid found in the shells of cashew nuts named anacardic acid. Later, the substance was found to consist of a family of related acid compounds(ref). It was used for many years for industrial purposes. “Anacardic acid is the main component of cashew nutshell liquid (CNSL), and finds use in the chemical industry for the production of cardanol, which is used for resins, coatings, and frictional materials. Cardanol is used to make phenalkamines, which are used as curing agents for the durable epoxy coatings used on concrete floors[9](ref). Life sciences research interest in anacardic acid it has been accelerating with the rise in knowledge about molecular biology and epigenetics. Once-unknown mechanisms underlying its antibacterial, anti-tumor, anti-inflammatory and other interesting properties are increasingly better understood, and the acid is of increasing interest for its practical anti-cancer and other health-inducing properties.
Anacardic acids occurs naturally in cashew nuts (the most accessible natural dietary source), mangos, and cashew apples. It is also found in Gingko biloba(ref), and geraniums.
The cashew nut developes in a shell which grows at the end of the cashew apple. Anacardic acid is mainly found in the shell.
Anacardic acid is a collective name. Chemists have identified dozens of molecular variants, The basic structure is this, where the tail element R is variable depending on the particular acid concerned: Image may be NSFW. Clik here to view.
Summing much of it up: “Anacardic acid, isolated from cashew shells or several other medicinal plants, is the general name given to a family of four different 6-alkyl salicyclic acids having varying degrees of unsaturation in the 15-carbon alkyl chain.1 These compounds are associated with anti-inflammatory, anti-tumor, molluscicidal, and anti-microbial activity. Literature frequently sites and gives the name anacardic acid to the completely-saturated compound (6-pentadecyl salicylic acid). Anacardic acid inhibits the histone acetyltransferase (HAT) activity of the transcription co-activators p300 and p300/CREB-binding protein-associated factor (pCAF) with IC50 values of 8.5 and 5 µM, respectively.2 At 25 µmol/L, anacardic acid suppresses NF-κB activation, inhibits IκB-α phosphorylation, and prohibits p65 nuclear translocation in KBM-5 cells.3(ref)”
“Diverse biological activities for the AAs have been described, including antimicrobial activity against methicillin-resistant bacteria [20–22], gastroprotection [23], and inhibition of the activity of several clinically targeted enzymes, such as lipoxygenase [24, 25], cyclooxygenase [26, 27], and histone acetyltransferases [28, 29]. It has been also demonstrated that AAs modulate the NF- B signaling pathway and inhibit tumor angiogenesis indicating that these compounds could be a therapeutic option in preventing or treating cancer [30–32](ref).”
Biological and health-related actions of anacardic acid
Like glucosamine and trehalose which we recently covered, there are many different molecular mechanisms and effects exercised by anacardic acids. Most interesting among many effects are the anti-inflammatory effects, the epigenetic effects and the antibacterial effects
Anti-inflammatory effects
Anacardic acid is an inhibitor of inflammation re suppression of expression of NF-kappaB. This is the start of the story – Anacardic acid can down regulate inflammation by inhibiting both the inducible and constitutive NF-kB. It also can reduce inflammation by inhibiting the lipoxygenases. This is probably due to HAT inhibition. Thus anacardic acid is a “epigenetic modulator of inflammation” by inhibiting the activation/expression of genes normally “turned on” by NF-kB. As you may know, TNF-alpha, IL-1beta, Lipopolysaccharide (LPS), PMA, OA, and EGF are main molecular activators of NF-kB. Anacardic acid inhibited the activation of NF-kB by all of these triggers of inflammation. This is why it appears to be an “upstream” inhibitor of NF-kB. Anacardic acid inhibited NF-kB induced gene expression in a dose-dependent relationship in vitro, with doses as low as 25 micro mol/L
The 2005 publication Lipoxygenase Inhibitory Activity of Anacardic Acidsreported: “6[8‘(Z)-Pentadecenyl]salicylic acid, otherwise known as anacardic acid (C15:1), inhibited the linoleic acid peroxidation catalyzed by soybean lipoxygenase-1 (EC 1.13.11.12, type 1) with an IC50 of 6.8 μM. The inhibition of the enzyme by anacardic acid (C15:1) is a slow and reversible reaction without residual activity. The inhibition kinetics analyzed by Dixon plots indicates that anacardic acid (C15:1) is a competitive inhibitor and the inhibition constant, KI, was obtained as 2.8 μM. Although anacardic acid (C15:1) inhibited the linoleic acid peroxidation without being oxidized, 6[8‘(Z),11‘(Z)-pentadecadienyl]salicylic acid, otherwise known as anacardic acid (C15:2), was dioxygenated at low concentrations as a substrate. In addition, anacardic acid (C15:2) was also found to exhibit time-dependent inhibition of lipoxygenase-1. The alk(en)yl side chain of anacardic acids is essential to elicit the inhibitory activity. However, the hydrophobic interaction alone is not enough because cardanol (C15:1), which possesses the same side chain as anacardic acid (C15:1), acted neither as a substrate nor as an inhibitor.”
The 2008 publication Anacardic acid (6-nonadecyl salicylic acid), an inhibitor of histone acetyltransferase, suppresses expression of nuclear factor-kappaB-regulated gene products involved in cell survival, proliferation, invasion, and inflammation through inhibition of the inhibitory subunit of nuclear factor-kappaBalpha kinase, leading to potentiation of apoptosisreported: “Anacardic acid (6-pentadecylsalicylic acid) is derived from traditional medicinal plants, such as cashew nuts, and has been linked to anticancer, anti-inflammatory, and radiosensitization activities through a mechanism that is not yet fully understood. Because of the role of nuclear factor-kappaB (NF-kappaB) activation in these cellular responses, we postulated that anacardic acid might interfere with this pathway. We found that this salicylic acid potentiated the apoptosis induced by cytokine and chemotherapeutic agents, which correlated with the down-regulation of various gene products that mediate proliferation (cyclin D1 and cyclooxygenase-2), survival (Bcl-2, Bcl-xL, cFLIP, cIAP-1, and survivin), invasion (matrix metalloproteinase-9 and intercellular adhesion molecule-1), and angiogenesis (vascular endothelial growth factor), all known to be regulated by the NF-kappaB. We found that anacardic acid inhibited both inducible and constitutive NF-kappaB activation; suppressed the activation of IkappaBalpha kinase that led to abrogation of phosphorylation and degradation of IkappaBalpha; inhibited acetylation and nuclear translocation of p65; and suppressed NF-kappaB-dependent reporter gene expression. Down-regulation of the p300 histone acetyltransferase gene by RNA interference abrogated the effect of anacardic acid on NF-kappaB suppression, suggesting the critical role of this enzyme. Overall, our results demonstrate a novel role for anacardic acid in potentially preventing or treating cancer through modulation of NF-kappaB signaling pathway.”
Epigenetic effects
Anacardic acid is a HAT (Histone Acetyl Transferase) inhibitor, meaning that it can help keep key genes such as those involved in inflammation, turned off. In fact, it is often used in laboratory experiments for that purpose. Specifically, Anacardic acid inhibits the Tip60 HAT as well as the p300/CBP HAT. Inhibiting the HAT p300/CBP is thought to be a great strategy for cancer treatment. p300/CBP can acetylate all four core histone proteins of the nucleosome on their lysine amino acids located on the histone protein tails. Both anacardic acid and curcumin and chemical variants of these have been investigated for their p300 HAT inhibiting capabilities.
The 2003 publication Small Molecule Modulators of Histone Acetyltransferase p300 reported:: “Histone acetyltransferases (HATs) are a group of enzymes that play a significant role in the regulation of gene expression. These enzymes covalently modify the N-terminal lysine residues of histones by the addition of acetyl groups from acetyl-CoA. Dysfunction of these enzymes is often associated with the manifestation of several diseases, predominantly cancer. Here we report that anacardic acid from cashew nut shell liquid is a potent inhibitor of p300 and p300/CBP-associated factor histone acetyltranferase activities. Although it does not affect DNA transcription, HAT-dependent transcription from a chromatin template was strongly inhibited by anacardic acid. Furthermore, we describe the design and synthesis of an amide derivative N-(4-chloro-3-trifluoromethyl-phenyl)-2-ethoxy-6-pentadecyl-benzamide (CTPB) using anacardic acid as a synthon, which remarkably activates p300 HAT activity but not that of p300/CBP-associated factor. Although CTPB does not affect DNA transcription, it enhances the p300 HAT-dependent transcriptional activation from in vitro assembled chromatin template. However, it has no effect on histone deacetylase activity. These compounds would be useful as biological switching molecules for probing into the role of p300 in transcriptional studies and may also be useful as new chemical entities for the development of anticancer drugs.”
The 2013 publication Anacardic acid, a histone acetyltransferase inhibitor, modulates LPS-induced IL-8 expression in a human alveolar epithelial cell line A549 reports: “Objective and design: The histone acetylation processes, which are believed to play a critical role in the regulation of many inflammatory genes, are reversible and regulated by histone acetyltransferases (HATs), which promote acetylation, and histone deacetylases (HDACs), which promote deacetylation. We studied the effects of lipopolysaccharide (LPS) on histone acetylation and its role in the regulation of interleukin (IL)-8 expression. Material: A human alveolar epithelial cell line A549 was used invitro. Methods: Histone H4 acetylation at the IL-8 promoter region was assessed by a chromatin immunoprecipitation (ChIP) assay. The expression and production of IL-8 were evaluated by quantitative polymerase chain reaction and specific immunoassay. Effects of a HDAC inhibitor, trichostatin A (TSA), and a HAT inhibitor, anacardic acid, were assessed. Results: Escherichia coli-derived LPS showed a dose- and time-dependent stimulatory effect on IL-8 protein production and mRNA expression in A549 cells in vitro. LPS showed a significant stimulatory effect on histone H4 acetylation at the IL-8 promoter region by ChIP assay. Pretreatment with TSA showed a dose-dependent stimulatory effect on IL-8 release from A549 cells as compared to LPS alone. Conversely, pretreatment with anacardic acid inhibited IL-8 production and expression in A549 cells. Conclusion: These data suggest that LPS-mediated proinflammatory responses in the lungs might be modulated via changing chromatin remodeling by HAT inhibition.”
Suppression of ROS-induced damage – Actions in mitochondria
Anacardic acids also are potent mitochondrial uncouplers. General anesthetics are also mitochondrial uncouplers which uncouple oxidative phosphorylation from the electron transport chain. This reduces ATP synthesis and also reduces ROS leak. Anacardic acid can uncouple oxidative phosphorylation from the electron transport chain. As a result, anacardic acid is a “mitochondrial-specific antioxidant” because they can change the membrane potential of the inner mitochondrial membrane, thereby reducing ROS leak.
Anacardic acid can exercise an uncoupling effect in liver mitochondria. The 2000 publication Uncoupling effect of anacardic acids from cashew nut shell oil on oxidative phosphorylation of rat liver mitochondriareports:“Anacardic acids are one of natural products found in not only the cashew nut shell oil but also the nut and fruit juice. The present study was conducted to investigate the uncoupling effect of anacardic acids on oxidative phosphorylation of rat liver mitochondria using succinate (plus rotenone) as a substrate. Four anacardic acids with C15:0, C15:1, C15:2 or C15:3 as an alkyl side chain exhibited uncoupling effects similar to the classical uncoupler, 2,4-dinitrophenol on ADP/O ratio, state 4 and respiratory control ratio (RCR). Anacardic acid with C15:1 side chain was most effective for uncoupling of these compounds. Salicylic acid, which has no alkyl side chain, exhibited a very weak uncoupling effect on oxidative phosphorylation. When the carboxyl group in anacardic acids was lost converting them to the corresponding cardanols, uncoupling activity dramatically decreased regardless of the number of double bonds in the long alkyl chain. These results suggest that the C15 alkyl side chain as well as the carboxyl group may play an important role in assisting the uncoupling activity of anacardic acids in liver mitochondria of animals. This study provides the first evidence of an uncoupling effect of anacardic acids on liver mitochondria.”
Anacardic acid can also help neutralize excess charge in liposomal membranes
The 2002 publication Anacardic acid-mediated changes in membrane potential and pH gradient across liposomal membranesreports: “We have previously shown that anacardic acid has an uncoupling effect on oxidative phosphorylation in rat liver mitochondria using succinate as a substrate (Life Sci. 66 (2000) 229–234). In the present study, for clarification of the physicochemical characteristics of anacardic acid, we used a cyanine dye (DiS-C3(5)) and 9-aminoacridine (9-AA) to determine changes of membrane potential (ΔΨ) and pH difference (ΔpH), respectively, in a liposome suspension in response to the addition of anacardic acid to the suspension. — These results provide the evidence for a unique function of anacardic acid, dissimilar to carbonylcyanide p-trifluoromethoxyphenylhydrazone or valinomycin, in that anacardic acid behaves as both an electrogenic (negative) charge carrier driven by ΔΨ, and a ‘proton carrier’ that dissipates the transmembrane proton gradient formed.” Assuming the same effect applies to mitochondrial membranes, then anacardic acid can help neutralize excess charge buildup and consequent ROS generation from old leaky mitochondria.
Regulating the DNA damage response in tumor cells
The2006 publication Inhibition of histone acetyltransferase activity by anacardic acid sensitizes tumor cells to ionizing radiation: reports: “Histone acetyltransferases (HATs) regulate transcription, chromatin structure and DNA repair. Here, we utilized a novel HAT inhibitor, anacardic acid, to examine the role of HATs in the DNA damage response. Anacardic acid inhibits the Tip60 HAT in vitro, and blocks the Tip60-dependent activation of the ATM and DNA–PKcs protein kinases by DNA damage in vivo. Further, anacardic acid sensitizes human tumor cells to the cytotoxic effects of ionizing radiation. These results demonstrate a central role for HATs such as Tip60 in regulating the DNA damage response. HAT inhibitors provide a novel therapeutic approach for increasing the sensitivity of tumors to radiation therapy.”
Because anacardic acid is a powerful HAT Inhibitor, it has many potential applications in medicine not only for cancer but for several other pathological conditions as well.
The 2009 publication Histone acetyl transferases as emerging drug targets reports: “Post-translational modifications, such as acetylation or phosphorylation, play a crucial role in the regulation of gene transcription in eukaryotes. Different subtypes of histone acetyl transferases (HATs) catalyze the acetylation of histones on specific lysine residues. A potential role of HATs in the pathology of cancer, asthma, COPD and viral infection has been described. This indicates that specific HAT inhibitors are potential tools for pharmacological research and might find therapeutic applications. This review focuses on the role of the HATs p300, CBP, PCAF and GCN5 in different diseases and the development of small-molecule inhibitors of these enzymes as potential drugs.”
Anti-cancer effects
It has been known for a long time that anacardic acid can act as a anticancer compound. See the 1993 publication Antitumor agents from the cashew (Anacardium occidentale) apple juice.And it can radiosensitize a tumor to external beam XRT. Many labs have already started synthesizing synthetic analogs of anacardic acid and are now trying them on cancer. Some of these anacardic acid analogs are already in clinical trials(ref).
The 2003 publication Small Molecule Modulators of Histone Acetyltransferase p3reported: “Histone acetyltransferases (HATs) are a group of enzymes that play a significant role in the regulation of gene expression. These enzymes covalently modify the N-terminal lysine residues of histones by the addition of acetyl groups from acetyl-CoA. Dysfunction of these enzymes is often associated with the manifestation of several diseases, predominantly cancer. Here we report that anacardic acid from cashew nut shell liquid is a potent inhibitor of p300 and p300/CBP-associated factor histone acetyltranferase activities. Although it does not affect DNA transcription, HAT-dependent transcription from a chromatin template was strongly inhibited by anacardic acid. Furthermore, we describe the design and synthesis of an amide derivative N-(4-chloro-3-trifluoromethyl-phenyl)-2-ethoxy-6-pentadecyl-benzamide (CTPB) using anacardic acid as a synthon, which remarkably activates p300 HAT activity but not that of p300/CBP-associated factor. Although CTPB does not affect DNA transcription, it enhances the p300 HAT-dependent transcriptional activation from in vitro assembled chromatin template. However, it has no effect on histone deacetylase activity. These compounds would be useful as biological switching molecules for probing into the role of p300 in transcriptional studies and may also be useful as new chemical entities for the development of anticancer drugs.”
The 2011 publication Anacardic acid inhibits estrogen receptor alpha-DNA binding and reduces target gene transcription and breast cancer cell proliferation discusses mechanicisms of action of anacardic acid in breast cancer cells beyond HAT inhibition, It relates: “Anacardic acid (2-hydroxy-6-alkylbenzoic acid) is a dietary and medicinal phytochemical with established anticancer activity in cell and animal models. The mechanisms by which anacardic acid inhibits cancer cell proliferation remain undefined. Anacardic acid 24:1ω5(AnAc 24:1ω5) was purified from geranium (Pelargonium × hortorum) and shown to inhibit the proliferation of estrogen receptor α (ERα)-positive MCF-7 and endocrine-resistant LCC9 and LY2 breast cancer cells with greater efficacy than ERα-negative primary human breast epithelial cells, MCF-10A normal breast epithelial cells, and MDA-MB-231 basal-like breast cancer cells. AnAc 24:1ω5 inhibited cell cycle progression and induced apoptosis in a cell-specific manner. AnAc 24:1ω5 inhibited estradiol (E2)-induced estrogen response element (ERE) reporter activity and transcription of the endogenous E2-target genes: pS2, cyclin D1, and cathepsin D in MCF-7 cells. AnAc 24:1ω5 did not compete with E2 for ERα or ERβ binding, nor did AnAc 24:1ω5 reduce ERα or ERβ steady state protein levels in MCF-7 cells; rather, AnAc 24:1ω5 inhibited ER-ERE binding in vitro. Virtual Screening with the molecular docking software Surflex evaluated AnAc 24:1ω5 interaction with ERα ligand binding and DNA binding domains (LBD and DBD) in conjunction with experimental validation. Molecular modeling revealed AnAc 24:1ω5 interaction with the ERα DBD but not the LBD. Chromatin immunoprecipitation (ChIP) experiments revealed that AnAc 24:1ω5 inhibited E2-ERα interaction with the endogenous pS2 gene promoter region containing an ERE. These data indicate that AnAc 24:1ω5 inhibits cell proliferation, cell cycle progression and apoptosis in an ER-dependent manner by reducing ER-DNA interaction and inhibiting ER-mediated transcriptional responses.”
A 2013 publication Inhibition of PCAF by anacardic acid derivative leads to apoptosis and breaks resistance to DNA damage in BCR-ABL-expressing cells: Acetylation of histones and nonhistone proteins is a posttranslational modification which plays a major role in the regulation of intracellular processes involved in tumorigenesis. It was shown that different acetylation of proteins correlates with development of leukemia. It is proposed that histone acetyltransferases (HATs) are important novel drug targets for leukemia treatment, however data are still not consistent. Our previous data showed that a derivative of anacardic acid - small molecule MG153, which has been designed and synthesized to optimize the HAT inhibitory potency of anacardic acid, is a potent inhibitor of p300/CBP associated factor (PCAF) acetyltransferase. Here we ask whether inhibition of PCAF acetyltransferase with MG153 will show proapoptotic effects in cells expressing BCR-ABL, which show increased PCAF expression and are resistant to apoptosis. We found that inhibition of PCAF decreases proliferation and induces apoptosis, which correlates with loss of the mitochondrial membrane potential and DNA fragmentation. Importantly, cells expressing BCR-ABL are more sensitive to PCAF inhibition compared to parental cells without BCRABL. Moreover, inhibition of PCAF in BCR-ABL-expressing cells breaks their resistance to DNA damage-induced cell death. These findings provide direct evidence that targeting the PCAF alone or in combination with DNA-damaging drugs shows cytotoxic effects and should be considered as a prospective therapeutic strategy in chronic myeloid leukemia cells. Moreover, we propose that anacardic acid derivative MG153 is a valuable agent and further studies validating its therapeutic relevance should be performed.
A study published last month (June 2014) Discovery of Protein Disulfide Isomerase P5 Inhibitors that Reduce the Secretion of MICA from Cancer Cellspoints to another anti-cancer property of anacardic acid: “In order to regulate the activity of P5, which is a member of the protein disulfide isomerase family, we screened a chemical compound library for P5-specific inhibitors, and identified two candidate compounds (anacardic acid and NSC74859). Interestingly,anacardic acid inhibited the reductase activity of P5, but did not inhibit the activity of protein disulfide isomerase (PDI), thiol-disulfide oxidoreductase ERp57, or thioredoxin. NSC74859 inhibited all these enzymes. When we examined the effects of these compounds on the secretion of soluble major histocompatibility complex class-I-related gene A (MICA) from cancer cells, anacardic acid was found to decrease secretion. In addition, anacardic acid was found to reduce the concentration of glutathione up-regulated by the anticancer drug 17-demethoxygeldanamycin in cancer cells. These results suggest that anacardic acid can both inhibit P5 reductase activity and decrease the secretion of soluble MICA from cancer cells. It might be a novel and potent anticancer treatment by targeting P5 on the surface of cancer cells.”
Anacardic acids inhibits the SUMO pathway in AML cancer cells enhancing chemotherapies.
Another last-month publication was The ROS/SUMO Axis Contributes to the Response of Acute Myeloid Leukemia Cells to Chemotherapeutic Drugs: “Chemotherapeutic drugs used in the treatment of acute myeloid leukemias (AMLs) are thought to induce cancer cell death through the generation of DNA double-strand breaks. Here, we report that one of their early effects is the loss of conjugation of the ubiquitin-like protein SUMO from its targets via reactive oxygen species (ROS)-dependent inhibition of the SUMO-conjugating enzymes. Desumoylation regulates the expression of specific genes, such as the proapoptotic gene DDIT3, and helps induce apoptosis in chemosensitive AMLs. In contrast, chemotherapeutics do not activate the ROS/SUMO axis in chemoresistant cells. However, pro-oxidants or inhibition of the SUMO pathway by anacardic acid restores DDIT3 expression and apoptosis in chemoresistant cell lines and patient samples, including leukemic stem cells. Finally, inhibition of the SUMO pathway decreases tumor growth in mice xenografted with AML cells. Thus, targeting the ROS/SUMO axis might constitute a therapeutic strategy for AML patients resistant to conventional chemotherapies.”
Endoplasmic reticulum stress and autophagy
Most of the properties of anacardic acid listed above have been known for years. However, there are also important new discoveries. As our knowledge of molecular biology and pathways involved in health and disease states become better known, we are discovering properties of the acid that were unknown before. For example, effective autophagyis now regarded to be an important tool for inducing death of some strains of cancer cells. The 2014 publication Induction of the endoplasmic reticulum stress and autophagy in human lung carcinoma A549 cells byanacardic acidreports: “Anacardic acid (AA, 2-hydroxy-6-pentadecylbenzoic acid), a constituent of the cashew-nut shell, has a variety of beneficial effects on the treatment of cancer and tumors. However, the fact that AA induces ER stress and autophagy in cancer cell is not known. We investigated the effect of AA on ER-stress and autophagy-induced cell death in cancer cells. Because of our interest in lung cancer, we used the non-small cell lung adenocarcinoma A549 cells treated with 3.0 μg/ml of AA for this research. In this research we found that AA induces intracellular Ca(2+) mobilization and ER stress. AA induced the ER stress-inducing factors, especially IRE1α, and the hallmarks of UPR, Grp78/Bip and GADD153/CHOP. AA inhibited the expression of p-PERK and its downstream substrate, p-elF2α. We also demonstrated that AA induces autophagy. Up-regulation of autophagy-related genes and the appearance of autophagosome in transfected cells with green fluorescent protein (GFP)-LC3 and GFP-Beclin1 plasmids showed the induction of autophagy in AA-treated A549 cells. The morphological analysis of intracellular organelles by TEM also showed the evidence that AA induces ER stress and autophagy. For the first time, our research showed that AA induces ER stress and autophagy in cancer cells.”
Mitochondrial-mediated apoptosis in cancer cells
The 2013 publication Anacardic acid induces mitochondrial-mediated apoptosis in the A549 human lung adenocarcinoma cells relates: “Anacardic acid (AA) is a constituent of the cashew nut shell and is known as an inhibitor of nuclear factor-κB (NF-κB). We investigated the cytotoxicity of AA on cancer cells and more experiments to reveal the cell death mechanism focused on A549 lung adenocarcinoma cells for our interest in lung cancer. To examine the molecular mechanism of cell death in AA treated A549 cells, we performed experiments such as transmission electron microscopy caspase-independent apoptosis with no inhibition of cytotoxicity by pan-caspase inhibitor, Z-VAD-fmk, in A549 cells. Our results showed the possibility of mitochondrial-mediated apoptosis through the activation of apoptosis-inducing factor (AIF) and an intrinsic pathway executioner such as cytochrome c. This study will be helpful in revealing the cell death mechanisms and in developing potential drugs for lung cancer using AA. (TEM), western blot analysis, fluorescence-activated cell sorting (FACS), genomic DNA extraction and staining with 4′,6-diamidino-2-phenylindole (DAPI). For the first time we revealed that AA induces “
Anacardic acid may enhance the proliferation of ovarian cancer cells
Surprise! There is another side to the cancer story. As is sometimes the case with health-producing substances, it appears that anacardic acid can be used by some cancer cells to enhance their survival and proliferation. The June 2014 publication Anacardic Acidenhances the proliferation of human ovarian cancer cellsreports: “BACKGROUND: Anacardic acid (AA) is a mixture of 2-hydroxy-6-alkylbenzoic acid homologs. Certain antitumor activities of AA have been reported in a variety of cancers. However, the function of AA in ovarian cancer, to date, has remained unknown. METHODS: Ovarian cancer cell lines were exposed to AA, after which cell proliferation, apoptosis, invasion and migration assays were performed. Phalloidin staining was used to observe lamellipodia formation. Reverse transcription polymerase chain reaction (RT-PCR) and western blotting were used to assess the mRNA and protein expression levels of Phosphatidylinositol 3-kinase (PI3K), vascular endothelial growth factor (VEGF) and caspase 3. RESULTS: Our results showed that AA promotes ovarian cancer cell proliferation, inhibits late apoptosis, and induces cell migration and invasion, as well as lamellipodia formation. AA exposure significantly up-regulated PI3K and VEGF mRNA and protein expression, while, in contrast, it down-regulated caspase 3 mRNA and protein expression in comparison to untreated control cells. CONCLUSION: Taken together, our results demonstrate for the first time that AA may potentiate the proliferation, invasion, metastasis and lamellipodia formation in ovarian cancer cell lines via PI3K, VEGF and caspase 3 pathways.”
I suspect that there might be a non-linear dose-response involved here. This was a cell-level study, and it is not clear what the implications are for in-vivo administration of anacardic acid. In any event, this new result clouds the picture when it comes to the impact of anacardic acid on cancers.
The 2011 publication Anacardic acid induces caspase-independent apoptosis and radiosensitizes pituitary adenoma cellsreports: “OBJECT: Pituitary adenomas, which are common intracranial tumors, are associated with significant patient morbidity due to hormone secretion or mass effect or as a complication of therapy. Epigenetic regulation has emerged as an important component of malignant tumor pathogenesis, although the contribution in the progression of benign pituitary tumors remains largely unexplored. The present study evaluates the effect of anacardic acid (6-pentadecyl salicylic acid), a natural histone acetyltransferase inhibitor, on pituitary adenoma cells. METHODS: The concentration- and time-dependent effects of anacardic acid on the viability of GH3 and MMQ pituitary adenoma cells were determined by 3-(4,5-dimethylthiazoyl-2-yl)-2,5-diphenyltetrazolium bromide assay. Cell cycle phase distribution, protein expression, and percentage of apoptotic cells were assessed by flow cytometry and Western blotting. Colony forming assays were used to study the radiosensitizing effect of anacardic acid. RESULTS: The present study identifies a novel antiproliferative and cytotoxic effect of anacardic acid on pituitary adenoma cells. These effects were associated with an increase in poly([adenosine diphosphate]-ribose) polymerase cleavage, sub-G1 arrest, and annexin V staining, consistent with apoptotic cell death; however, the pancaspase inhibitor carbobenzoxy-valyl-alanyl-aspartyl-(O-methyl)-fluoromethylketone failed to reverse anacardic acid-induced cell death, suggesting a possible nonclassical apoptotic mechanism. Anacardic acid also reduced the expression of survivin and X-linked inhibitor of apoptosis protein, antiapoptotic proteins associated with cellular survival and radioresistance, and radiosensitized pituitary adenoma cells. CONCLUSIONS: These findings warrant further exploration of anacardic acid as a single agent or as an adjunct to radiation therapy for the treatment of pituitary tumors.”
Induction of developmental changes in global gene expression during developmental stages
This was demonstrated in the Plasmodium falciparum parasite, the one that causes malaria. The 2008 publication Histone acetyltransferasere inhibitor Anacardic Acid Causes Changes in Global Gene Expression during In Vitro Plasmodium falciparum Development: “To better understand the role of histone lysine acetylation in transcription in Plasmodium falciparum, we sought to attenuate histone acetyltransferase (HAT) activity using anacardic acid (AA). We showed that AA reversibly and noncompetitively inhibited the HAT activity of recombinant PfGCN5. To a lesser extent, AA inhibited the PfGCN5 activity in parasite nuclear extracts but did not affect histone deacetylase activity. AA blocked the growth of both chloroquine-sensitive and -resistant strains, with a 50% inhibitory concentration of ∼30 μM. Treatment of the parasites with 20 μM of AA for 12 h had no obvious effect on parasite growth or gross morphology but induced hypoacetylation of histone H3 at K9 and K14, but not H4 at K5, K8, K12, and K16, suggesting inhibition of the PfGCN5 HAT. Microarray analysis showed that this AA treatment resulted in twofold or greater change in the expression of 271 (∼5%) parasite genes in late trophozoites, among which 207 genes were downregulated. Cluster analysis of gene expression indicated that AA mostly downregulated active genes, and this gene pool significantly overlapped with that enriched for H3K9 acetylation. We further demonstrated by chromatin immunoprecipitation and real-time PCR that AA treatment reduced acetylation near the putative promoters of a set of downregulated genes. This study suggests that the parasiticidal effect of AA is at least partially associated with its inhibition of PfGCN5 HAT, resulting in the disturbance of the transcription program in the parasites.”
Anti-microbial and anti-fungal activities
Anacardic acid has wonderful antibacterial and anti fungal properties that have been known since 1969. It can kill helicobacter pylori in the stomach and heal gastritis or gastric ulcers caused by this bacteria. The most likely antibiotic mechanism of action is the fact that anacardic acid can act as an iontophore, an electrically charged molecule. See Selective ionophoric properties of anacardic acid (1995) It is a very good anti fungal as well.
Anacardic acid is effective against MRSA bacteria
Probably the most interesting application of anacardic acid is that it is very effective for resistant strains of Staph aureus (MRSA). These are staph bacteria that have evolved to be resistant to almost all conventional bacteria creating a significant threat to public health(ref). The 2003 publication Antibacterial Action of Anacardic Acids against Methicillin ResistantStaphylococcus aureus (MRSA)reports “The structural and antibacterial activity relationship of 6-alk(en)ylsalicylic acids, also known as anacardic acids, was investigated against Gram-positive bacteria, emphasizing the methicillin resistant Staphylococcus aureus ATCC 33591 (MRSA) strain. The unsaturation in the alkyl side chain is not essential in eliciting activity but is associated with increasing the activity. The antibacterial activity of methicillin against MRSA strains was significantly enhanced in combination with C12:0-anacardic acid, and the fractional inhibitory concentration index for this combination was calculated as 0.281. It appears that biophysical disruption of the membrane (surfactant property) is due to the primary response to their antibacterial activity, while biochemical mechanisms are little involved. The compounds possessing the similar log Pvalues exhibit similar activity.”
sensing-controlled virulence factors of Chromobacterium violaceum and Pseudomonas aeruginosa reports: “BACKGROUND AND AIMS: Quorum sensing (QS) is a process of bacterial cell-cell communication that controls a large number of systems affecting pathogenicity. Interrupting this communication system can provide nonvirulent pathogenic bacteria. The aim of this study was to evaluate the anti-quorum sensing (anti-QS) potential of an anacardic acids mixture isolated from Amphipterygium adstringens, a medicinal plant known as “cuachalalate”, to prevent the onset of bacterial infections as an alternate to antibiotics. METHODS: Initially we investigated the anti-QS activity of A. adstringens hexane extract (HE) by the inhibition of violacein production in Chromobacterium violaceum. From the active HE, an anacardic acid mixture (AAM) was obtained. The anti-quorum sensing activity of AAM was investigated by the rhamnolipid and pyocyanin production constraint as well as decrease of elastase activity, all being quorum sensing-controlled virulence factors expressed in the pathogenic bacteria Pseudomonas aeruginosa. RESULTS: HE induced a 91.6% of inhibition of the violecin production at 55 μg/mL concentration, whereas AAM showed 94% of inhibition at 166 μg/mL. In both cases, inhibition of violacein production did not affect the viability of the bacterium. AAM inhibited pyocyanin (86% at 200 μg/mL) and rhamnolipid (91% at 500 μg/mL) production in a dose/response form and decrease the elastase (75% at 500 μg/mL) activity in P. aeruginosa without affecting its development. CONCLUSIONS: Because an anacardic acids mixture isolated from A. adstringens demonstrated anti-QS, it could be further exploited for novel molecules to treat the emerging infections of antibiotic-resistant bacterial pathogens.”
Use in dentistry
Anacardic acid derivatives might be useful for the prevention of dental cavities. The publication Design and evaluation of anacardic acid derivatives as anticavity agentsreports: “On the basis of antibacterial anacardic acids, 6-pentadecenylsalicylic acids, isolated from the cashew apple, Anacardium occidentale L. (Anacardiaceae), a series of 6-alk(en)ylsalicylic acids were synthesized and tested for their antibacterial activity against Streptococcus mutans ATCC 25175. – Among them, 6-(4′,8′-dimethylnonyl)salicylic acid was found to exhibit the most potent antibacterial activity against this cariogenic bacterium with the minimum inhibition concentration (MIC) of 0.78 μg/ml.” The antibacterial activity is probably associated with suppression of quorum sensing and bacterial colony formation.
Anti-gout properties
Anacardic acid inhibits xanthine oxidase, an important enzyme in the formation of uric acid. Gout is a condition where uric acid accumulates and precipitates in joints and in soft tissue, causing gouty arthritis. Hypoxanthine is converted into xanthine, which is then converted into uric acid. The enzyme that catalyzes the conversion of hypoxanthine to xanthine and then to uric acid is called Xanthine oxidase. Anacardic acid inhibits this enzyme.
The 2003 publication Characterization of xanthine oxidase inhibition by anacardic acidsrelates: “Anacardic acid, 6[8(Z), 11(Z), 14-pentadecatrienyl]salicylic acid, inhibits generation of superoxide radicals by xanthine oxidase. This inhibition does not follow a hyperbolic inhibition, depends on anacardic acid concentrations, but follows a sigmoidal inhibition. — The results indicate that anacardic acid binds to allosteric sites near the xanthine-binding domain in xanthine oxidase. Salicylic acid moiety and alkenyl side chain in anacardic acid are associated with the cooperative inhibition and hydrophobic binding, respectively.”
Anacardic acid may help protect lungs from air pollutants
The 2013 publication Anacardic Acids from Cashew Nuts Ameliorate Lung Damage Induced by Exposure to Diesel Exhaust Particles in Micereports: “Anacardic acids from cashew nut shell liquid, a Brazilian natural substance, have antimicrobial and antioxidant activities and modulate immune responses and angiogenesis. As inflammatory lung diseases have been correlated to environmental pollutants exposure and no reports addressing the effects of dietary supplementation with anacardic acids on lung inflammation in vivo have been evidenced, we investigated the effects of supplementation with anacardic acids in a model of diesel exhaust particle- (DEP-) induced lung inflammation. BALB/c mice received an intranasal instillation of 50 μg of DEP for 20 days. Ten days prior to DEP instillation, animals were pretreated orally with 50, 150, or 250 mg/kg of anacardic acids or vehicle (100 μL of cashew nut oil) for 30 days. The biomarkers of inflammatory and antioxidant responses in the alveolar parenchyma, bronchoalveolar lavage fluid (BALF), and pulmonary vessels were investigated. All doses of anacardic acids ameliorated antioxidant enzyme activities and decreased vascular adhesion molecule in vessels. Animals that received 50 mg/kg of anacardic acids showed decreased levels of neutrophils and tumor necrosis factor in the lungs and BALF, respectively. In summary, we demonstrated that AAs supplementation has a potential protective role on oxidative and inflammatory mechanisms in the lungs.”
Anacardic acid might be useful for reducing pain hypersensitivity after surgical incisions.
The regulation of gene expression in nociceptive pathways contributes to the induction and maintenance of pain sensitization. Histone acetylation is a key epigenetic mechanism controlling chromatin structure and gene expression. Chemokine CC motif receptor 2 (CXCR2) is a proinflammatory receptor implicated in neuropathic and inflammatory pain and is known to be regulated by histone acetylation in some settings. The authors sought to investigate the role of histone acetylation on spinal CXCR2 signaling after incision. METHODS: Groups of 5-8 mice underwent hind paw incision. Suberoylanilide hydroxamic acid and anacardic acid were used to inhibit histone deacetylase and histone acetyltransferase, respectively. Behavioral measures of thermal and mechanical sensitization as well as hyperalgesic priming were used. Both message RNA quantification and chromatin immunoprecipitation analysis were used to study the regulation of CXCR2 and ligand expression. Finally, the selective CXCR2 antagonist SB225002 was administered intrathecally to reveal the function of spinal CXCR2 receptors after hind paw incision. RESULTS: Suberoylanilide hydroxamic acid significantly exacerbated mechanical sensitization after incision. Conversely, anacardic acid reduced incisional sensitization and also attenuated incision-induced hyperalgesic priming. Overall, acetylated histone H3 at lysine 9 was increased in spinal cord tissues after incision, and enhanced association of acetylated histone H3 at lysine 9 with the promoter regions of CXCR2 and keratinocyte-derived chemokine (CXCL1) was observed as well. Blocking CXCR2 reversed mechanical hypersensitivity after hind paw incision.”
Anacardic acid is closely related to urushiol, the active toxin in poison ivy, poison oak and poison sumac – and exposing the skin to it may lead to contact dermatitis
Here is another down side, The 2012 publication Urushiol-induced contact dermatitis caused during Shodhana (purificatory measures) of Bhallataka (Semecarpus anacardium Linn.) fruitrelates: “Bhallataka (Semecarpus anacardium Linn.; Ancardiaceae) is mentioned under Upavisha group in Ayurvedic classics and it is described as a poisonous medicinal plant in Drugs and Cosmetics Act (India), 1940. Fruit of Bhallataka is used either as a single drug or as an ingredient in many compound formulations of Indian systems of medicine to cure many diseases. Tarry oil present in the pericarp of the fruit causes blisters on contact. The major constituent of the tarry oil is anacardic acid and bhilawanol, a mixture of 3-n-pentadec(en)yl catechols. Bhilawanol A and B are known as Urushiols, and also, anacardic acid is closely related to Urushiol. Urushiol-induced contact dermatitis is the medical name given to allergic rashes produced by the oil Urushiol. This paper deals with five case reports of contact dermatitis caused during different stages of Shodhana (purificatory measures) of Bhallataka fruit due to improper handling of the utensils and disposal of media used in Shodhana procedure and their Ayurvedic management. 00.” If you eat a cashew nut raw, it can burn your mouth!
Pharmaceutical companies have been exploring variants of Anacardic acid for the development of drugs.
As is often the case when there is a plant-based substance that has important biological effects, drug companies have been looking for biological analogs. The “good” reasons usually cited are that a substance is wanted with more powerful biological effects, better bioavailability and less side effects. Skeptics point out that often the “real” reason is that a molecule is wanted that can be patented and owned. No drug company is ready to spring for hundreds of millions of dollars for a clinical trial for an unpatentable natural substance. “There ain’t no money in something that grows in a plant and that anybody can sell cheap.” So, some of the studies mentioned are based on testing chemical analogs.
The 2010 publication Improved inhibition of the histone acetyltransferase PCAF by an anacardic acid derivativerelates: “Several lines of evidence indicate that histone acetyltransferases (HATs) are novel drug targets for treatment of diseases like, for example, cancer and inflammation. The natural product anacardic acid is a starting point for development of small molecule inhibitors of the histone acetyltransferase (HAT) p300/CBP associated factor (PCAF). In order to optimize the inhibitory potency, a binding model for PCAF inhibition by anacardic acid was proposed and new anacardic acid derivatives were designed. Ten new derivatives were synthesized using a novel synthetic route. One compound showed a twofold improved inhibitory potency for the PCAF HAT activity and a twofold improved inhibition of histone acetylation in HEP G2 cells.”
The 2007 publication Molecular design of anti-MRSA agents based on the anacardic acid scaffolddescribes such a search for analogs for anacardic acid: “A series of anacardic acid analogues possessing different side chains viz. phenolic, branched, and alicyclic were synthesized and their antibacterial activity tested against methicillin-resistant Staphylococcus aureus(MRSA). The maximum activity against this bacterium occurred with the branched side-chain analogue, 6-(4′,8′-dimethylnonyl)salicylic acid, and the alicyclic side-chain analogue, 6-cyclododecylmethyl salicylic acid, with the minimum inhibitory concentration (MIC) of 0.39 μg/mL, respectively. This activity was superior to that of the most potent antibacterial anacardic acid isolated from the cashew Anacardium occidentale (Anacardiaceae), apple and nut, that is, the 6-[8′(Z),11′(Z),14′-pentadecatrienyl]salicylic acid.” Also, there is the 2013 publication Efficient synthesis ofanacardic acidanalogues and their antibacterial activities.
Looking forward
The molecular actions and health-producing properties of anacardic acid documented here seem amazing, but there appears to be a lot more we don’t know about in-vivo actions of the substance. For example, after building a very convincing case for its anti-cancer activities above, we cited a publication that indicates that anacardic acid may enhance the “proliferation, invasion, metastasis and lamellipodia formation in ovarian cancer cell lines.” What gives? I expect we will continue to hear more about it as time goes on.
Plants are far from being deaf and dumb. We think this blog may help convince you that they are smart, listen carefully and are highly expressive.
The topic of this blog is plant communication – communications within different parts of one plant, between nearby plants of the same species, between plants of different species they intersect with, between plants and predatory insects that will eat herbivores that are destroying the plant, and multi-way communications between plants and insects and microbial organsms that can help a plant deal with stresses and counter predators. We have much In common with plants, sharing about a quarter of our DNA with them and utilizing many of the biological pathways they use. Besides being fascinating, understanding plants may help us in many ways including how to prevent or limit cancers and other diseases, and how to live long healthy lives. Plant signaling may have strong implications for creating sustainable agriculture, for identifying and understanding health-producing phyto-substances and even for gardening and getting on better with our house plants. Image may be NSFW. Clik here to view.Image source
We first became interested in plant communications after reading well-known biologist Daniel Chamovitz’s book What a Plant Knows. Chamovitz is Director of the Manna Center for Plant Biosciencesat Tel Aviv University). HIs lab is focused primarily on understanding the role of the COP9 signalosome (CSN) in regulating development. The CSN functions at the nexus of signaling inputs and ubiquitin-mediated protein degradation pathways. The lab is using a combination of genetic, molecular and genomic approaches to dissect the varied roles of the CSN in the development of plants and animals, using both Arabidopsis and Drosophila as model systems.
But we will not be getting into such detailed matters here. Readers of this blog know we often love to dig into complex molecular biology pathways. We don’t do that in this blog entry because there are several basics to be told first, and because those pathways in plants are less explored as compared to in animals. Because of the rich variety of plant communications and depth of this field, we necessarily confine ourselves in this introductory blog post to illustrative examples, leaving out much interesting detail.
Why plant communications in a longevity science blog?
Plant signaling is important for humans and for their defenses and longevity. Plant signaling is a defense mechanism for the plants immune system, that is, a system that detects dangers to the wellbeing of an organism and takes defensive actions. Plant communications can warn other parts of the same plant, other plans of the same species, or other nearby plants of different species that there is an invader coming to harm them. This allows a multiplicity of defensive action to be taken. Because plant immunity communications systems work very differently from ours, understanding how they work may facilitate us to augment our own immunity responses and thus contribute to our longevity.
Plants have much in common with us and can exhibit remarkable behavior once thought to be possible only in animals with nervous systems and brains. Plant communications are part of plant behaviors – what plants pro-actively do. These behaviors are varied and complex, despite our perceiving plants just sitting there passively and accepting what comes. Plants behave in respond to light and shade, to the availability of water and nutrients, to heat and cold, to seasons, to what is in their neighborhood, to changes in air composition, to other stresses and to the presence of specific predators. They have memory and predictive capabilities. They can signal to warn themselves of a danger, to warn neighboring plants, to organize individual and collective defenses, and to enroll members of other species that can help them in an emergency. Plants live together in a very complicated and interconnected ecological system. They talk back and forth with many other species. The more we can understand about the communications mechanisms, the more the possibilities for the development of sustainable applications to agricultural and the more we can possibly learn about other species, including ourselves.
The purpose of plant communications, as much as has been established so far by science, is plant development and survival on multiple levels – for individual plants, for communities of plants and for species. It is a defensive mechanism mainly initiated by experienced stresses or survival needs. One important form of signaling we will discuss is release of volatile organic compounds (VOCs), chemical cues given off by a damaged plant. But, as we will see, plants can communicate by counterparts of all of our own sensory modalities.
What this blog entry is not about
There are many forms of molecular plant communications that happen in cell-level and intercellular signaling pathways, just like in bacterial, animal and all other life forms. We mention there is an important class of internal signaling molecules, phytohormones. Phytohormones regulate all the stages of a plants lifecycle and its responses to stresses. A PowerPoint presentation on phytohormones can be downloaded from this site. Two of the phytohormone families are stress responsive and of particular relevance in this regard, Salicylates and Jasmonates Further there is much important internal signaling that takes place via the phloem, an important vascular system in plants. The phloem is made up of live cells – active tubes that transport nutrients, particularly sugars produced by photosynthesis in the leaves. In trees it is the innermost layer of the bark. The phloem as a signaling conduit is covered in the 2005 publication Multitude of Long-Distance Signal Molecules Acting Via Phloem. Also, plants like us humans can utilize microRNAs for inter-cellular and inter organ communications. See “And yet it moves”: Cell-to-cell and long-distance signaling by plant microRNAs. Interesting though these topics are, in this blog entry we will not focus on such basically internal plant signaling pathways. Instead, we focus on forms of communications capable of reaching other plants or organisms belonging to other species.
Plant communication was all the rage in the early 1980’s with the “talking trees” theory popularized in the media. But then the idea was quickly discredited in scientific circles in reaction to several studies that could not be replicated. However, by the late 1990’s a new group of verifiable studies began to emerge re-enforcing the existence of complex signaling process among plants. This blog will present a number of ways that plant signalling/communication/eavesdropping has been documented.
Plants communicate via the same senses humans and animals do, using visual, sound, touch and smell signaling
For communications purposes we may use visual clues, sound, touch, and smell, our communications being based on specialized organs optimized for our survival. Plants too sense and communicate via visual clues, sound, touch and smell – but using completely different perceptive and information processing means. We also communicate internally via electrical signaling, such as at synaptic junctions. Plants also utilize electrical signaling. We provide selected examples of each modality. Image may be NSFW. Clik here to view.Image source
Visual communications
It has been well accepted for some time that Plants reprogram their growth responding to light and shade.Everybody who tends to plants knows this. What they may not know is that plants can predict when shade can be expected from other plants and proactively take actions to keep themselves in the light.
The 1999 publication Keeping up with the neighbours: phytochrome sensing and other signalling mechanisms reported: “Plants ‘forage’ for light in plant canopies using a variety of photosensory systems. Far-red radiation (FR) reflected by neighbours is an early signal of competition that elicits anticipatory shade-avoidance responses. In Arabidopsis and cucumber, perception of reflected FR requires phytochrome B. Horizontal blue (B) light gradients also guide plant shoots to canopy gaps in patchy vegetation, and these B light signals are perceived by specific photoreceptors. When plants are shaded by neighbours they undergo extensive reprogramming of their morphological development. Although phytochromes and B light receptors are certainly involved in these responses to shading, other sensory systems probably play important roles in the field. Recent studies of plant–plant signalling are unveiling a paradigm of sensory diversity and sophistication, which has important implications for understanding the functioning of plant populations and communities.” So, the implicit visual message put out by a plant shooting our new foliage is “Sorry friends, but I have to spread my leaves.” The message received by plants potentially being cast into shade is “Some other plant or plants could be putting me in the shade; I have to do something about that.” The plant response could be long shoots, favoring growth on one side, leaf adaption, etc.
Looking out our office window, we can see semi-shaded patches of wild vegetation, trees, bushes, vines, leafy plants and grasses – sometimes dozens or hundreds of plant species in just a tiny area. We have often wondered how equilibrium could be established involving so many different proximate species. We speculate that this is a result of different species employing somewhat different survival approaches. It all seems very tranquil, but actually there is probably a lot of inter-species arguing, shouting and even screaming going on. The distribution of roots, branches, shoots and leaves is in large measure the collective result of the competitive resource-seeking efforts of all those plants. All that greenery seems to grow naturally and peacefully, but what is happening is an intense multi-way game of anticipatory and responsive communication, competition, and sometimes cooperation in interaction with other species such as fungi and insects. A major resource being light, it seems that trees have a big advantage if and when they can manage to grow up.
Touch communications
It has long been known that simply touching the leaves of some plants can initiate behavioral responses such as leaf curling. If an insect touches two hairs within 20 seconds on the inner surface of a venus flytrap plant, the plant will instantly spring shut entrapping the insect so it can be digested by the plant. There are a great many other examples of “smart” plant sensitivity to touch
The 1990 publication Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis reports: “In response to water spray, subirrigation, wind, touch, wounding, or darkness, Arabidopsis regulates the expression of at least four touch-induced (TCH) genes. Ten to thirty minutes after stimulation, mRNA levels increase up to 100-fold. Arabidopsis plants stimulated by touch develop shorter petioles and bolts. This developmental response is known as thigmomorphogenesis. TCH 1 cDNA encodes the putative Arabidopsis calmodulin differing in one amino acid from wheat calmodulin. Sequenced regions of TCH 2 and TCH 3 contain 44% and 70% amino acid identities to calmodulin, respectively. The regulation of this calmodulin-related gene family in Arabidopsis suggests that calcium ions and calmodulin are involved in transduction of signals from the environment, enabling plants to sense and respond to environmental changes.” See also the 1993 publication Thigmomorphogenesis: the effect of mechanical perturbation on plants.
From The Myth of Stoic Trees: “Plants, being immobile, have responses to their environment often quite different from organisms that can escape unsuitable conditions. Therefore, plant responses to touch (often termed mechanical perturbation or MP in the scientific literature) can be exquisitely sensitive. The ability of some carnivorous plants to actively trap food is an example of touch response, as is leaf movement of sensitive plants (Mimosa spp.), and coiling of vine tendrils. == Thigmomorphogenesis can be induced by many types of environmental MP including wind, water spray, snow load, and rubbing from other plants. People, wild and domesticated animals, and even insects can also cause these changes. The responses are species-specific in terms of the amount of MP required and in the morphological changes seen. Initially studied in annual crop plants, such as peas, beans, corn, and sunflowers, MP was universally seen to decrease stem elongation and increase stem thickness. Other characteristics include shorter petiole length, decreased needle elongation, smaller leaves, reduced flower number, and increased senescence (programmed tissue death). Similar responses have been demonstrated in woody species including pine (Pinus), spruce (Picea), fir (Abies), poplar (Populus), and elm (Ulmus). Continual rubbing or brushing of woody trees and shrubs, even that which is gentle enough not to abrade tissue, will result in shorter heights and wider trunks. This is partially meditated through the release of ethylene gas, a naturally produced plant growth regulator, which in turn increases the formation of lignin in the disturbed tissues. The result of thigmomorphogenesis is a stocky, sturdy plant that is more resistant to breakage or windthrow than one that has been untouched, and the greater the disturbance the more pronounced the response. The short, stunted appearance of alpine forest trees is an extreme example of wind-induced thigmomorphogenesis. Such trees are less likely to break from snowload or suffer windthrow than thin, upright specimens.”
So a tree can respond to a massage. But this is not a New Age blog; we are interested in effects documented by hard scientific research.
The 2005 publication In touch: plant responses to mechanical stimuli reports: “Perception and response to mechanical stimuli are likely essential at the cellular and organismal levels. Elaborate and impressive touch responses of plants capture the imagination as such behaviors are unexpected in otherwise often quiescent creatures. Touch responses can turn plants into aggressors against animals, trapping and devouring them, and enable flowers to be active in ensuring crosspollination and shoots to climb to sunlit heights. Morphogenesis is also influenced by mechanical perturbations, including both dynamic environmental stimuli, such as wind, and constant forces, such as gravity. Even individual cells must sense turgor and wall integrity, and subcellular organelles can translocate in response to mechanical perturbations. Signaling molecules and hormones, including intracellular calcium, reactive oxygen species, octadecanoids and ethylene, have been implicated in touch responses. Remarkably, touch-induced gene expression is widespread; more than 2.5% of Arabidopsis genes are rapidly up-regulated in touch-stimulated plants. Many of these genes encode calcium-binding, cell wall modifying, defense, transcription factor and kinase proteins. With these genes as tools, molecular genetic methods may enable elucidation of mechanisms of touch perception, signal transduction and response regulation.”
The 2012 publication Arabidopsis touch-induced morphogenesis is jasmonate mediated and protects against pests reports: “Plants cannot change location to escape stressful environments. Therefore, plants evolved to respond and acclimate to diverse stimuli, including the seemingly innocuous touch stimulus [1-4]. Although some species, such as Venus flytrap, have fast touch responses, most plants display more gradual touch-induced morphological alterations, called thigmomorphogenesis [2, 3, 5, 6]. Thigmomorphogenesis may be adaptive; trees subjected to winds develop less elongated and thicker trunks and thus are less likely damaged by powerful wind gusts [7]. Despite the widespread relevance of thigmomorphogenesis, the regulation that underlies plant mechanostimulus-induced morphological responses remains largely unknown. Furthermore, whether thigmomorphogenesis confers additional advantage is not fully understood. — Here we show that jasmonate (JA) phytohormone both is required for and promotes the salient characteristics of thigmomorphogenesis in Arabidopsis, including a touch-induced delay in flowering and rosette diameter reduction. Furthermore, we find that repetitive mechanostimulation enhances Arabidopsis pest resistance in a JA-dependent manner. These results highlight an important role for JA in mediating mechanostimulus-induced plant developmental responses and resultant cross-protection against biotic stress.”
Smell communications
A central focus here is on molecular be communications via VOCs (volatile organic compounds) – giving off odors and smelling in animal terms. This is an area where plants do a better job than humans based on their millions of years of evolution, a very essential communications means for plants. These VOCs can inform other parts of a plant or other nearby plants to respond/emit chemicals that ward off attackers/curl leaves etc to repel the attacker. The distance VOCs travel, for the most part is 60cm or less though some only travel 15cm. It is not so well established if the intention is only to inform nearby plants of the same species or not. But there is often a defense response by other plant species if they are close enough. First, we will frame this branch of plant science research in a historical context.
The double about-face of science about plant smell communications
There were publications in the 1980s that plants communicate via volatile substances with each other about threats of being eaten. These publications were flawed in their research methodology and were soon discredited by publications from reputable scientists. However, evidence for such effects continued to accumulate and now there is little doubt that such communication exists. Plants of a number of species receiving communications from plants threatened or under attack mount defenses and are less likely to be eaten.
The 2014 publication Volatile communication between plants that affects herbivory: a meta-analysis Ecology Letters reviews the situation with respect to VOC signaling caused by herbivore attacks, i.e. signaling from the plant when it is being eaten or threatened to be eaten. The publication states in summary “Volatile communication between plants causing enhanced defense has been controversial. Early studies were not replicated, and influential reviews questioned the validity of the phenomenon. We collected 48 well-replicated studies and found overall support for the hypothesis that resistance increased for individuals with damaged neighbours. Laboratory or greenhouse studies and those conducted on agricultural crops showed stronger induced resistance than field studies on undomesticated species, presumably because other variation had been reduced. A cumulative analysis revealed that early, non-replicated studies were more variable and showed less evidence for communication. — These results indicate that plants of diverse taxonomic affinities and ecological conditions become more resistant to herbivores when exposed to volatiles from damaged neighbours.”
First statements of the volatile communications hypothesis
Going on: “When some plants are attacked by herbivores, they release chemical cues that cause other individuals to change their traits and become more resistant to herbivory. Communication between plants was first observed and reported more than 30 years ago (Rhoades 1983; Baldwin & Schultz 1983) and the number of reported cases has grown rapidly in the recent past (Heil & Karban 2010). We consider a process to be plant volatile communication if it involves signalling by a plant that causes a response in the same or a different individual that receives the cue. Emission or display of a cue is plastic and the response of the receiver is conditional on receiving the cue. We require that emitting the cue could potentially benefit the emitter, although this has proven difficult or impossible to establish (Karban 2008). For an interaction to be considered communication, the responder must have the choice of responding to the cue or not, a requirement which excludes allelopathy (Schenk & Seabloom 2010). We make no assumptions about the intended target of the cues; many plants use volatile cues to co-ordinate their own defences against herbivores when one branch is attacked and other branches on the same individual respond by increasing defences (Karban et al. 2006; Frost et al. 2007; Heil & Silva Bueno 2007; Rodriguez-Saona et al. 2009). There is no agreed upon definition of communication; our use of the term is broader than most and includes phenomena that some authors prefer to call eavesdropping or signalling. — Early reports of plant communication met with great interest from scientists and the popular press. David Rhoades observed that caterpillars placed on willow trees near damaged neighbours grew less well than those placed on trees near undamaged neighbours (Rhoades 1983). He hypothesised that the reduction in performance was caused by airborne communication from the damaged trees that increased resistance in neighbours. However, he was unable to repeat his initial results and his experimental design lacked true replication (D. Rhoades, pers. comm., Fowler & Lawton 1985). In addition, the poor performance of caterpillars on trees in close proximity to infested neighbours could have been caused by the introduction of insect pathogens rather than by communication between plants. Early experiments conducted on plants in growth chambers reported that plants exposed to volatiles coming from chambers containing feeding herbivores became more resistant, but the experimental designs used in these studies also lacked true replication (Baldwin & Schultz 1983; Bruin et al. 1992).”
The first about-face
“Following an influential review by Simon Fowler and John Lawton, most ecologists decided that communication between plants was a phenomenon that had been considered and debunked and that the phenomenon did not occur in nature (Fowler & Lawton 1985; Dicke & Bruin 2001). In addition to the limitations associated with experimental design of the early studies, communication between plants that benefited neighbours did not make sense to many ecologists. Early descriptions of this phenomenon were referred to as ‘talking trees’ by both the popular press and some scientists in the field. Natural selection would not be expected to favour the emission of cues that provided neighbouring competitors with information about herbivores.”
The second about-face
“Coinciding with the resurgence in scientific interest in communication has been a resolution of this apparent contradiction. — Many studies involving diverse plants reported evidence of volatile communication resulting in increased resistance to herbivore attack, indicating that this is a widespread natural phenomenon. Unlike early studies of this phenomenon, the studies considered in this review were well-replicated with independent sampling units. — Many of these studies did not identify mechanisms involved, even whether a plant response was responsible for the effects. Alternative hypotheses, such as direct repellency of herbivores by volatiles, were often not ruled out and would be worth considering in future studies. Determining the plant responses involved, particularly the volatile cues that were responsible will be well worth future effort. As expected, conditions that minimised background variation, particularly laboratory studies and studies of genetically homogeneous crop species, were more likely to detect significant effects of volatile cues on induced resistance. Future studies are needed to separate effects due to experimental conditions (laboratory vs. field) from those caused by different inducing agents (herbivores vs. artificial damage”
Up to this point in our discussion: 1. We know that communications among plants warning of herbivore attacks must exist because plants that are neighbors of attacked plants are less damaged in subsequent attacks, 2. With high probability VOCs are involved, 3. The exact mechanisms of initiation of communications, forms of protection and mechanisms of protection are less studied. We go on to discuss these issues.
Plant VOC communications in response to predatory attacks can be highly specific and can involve multiple kinds of VOCs including release of 1. substances that are repellant to the specific predator, 2. substances that are toxic to the specific predator, 3 substances that are alerts to other parts of the same plant or neighboring plants that tell them to upgrade a kind of predator defense, and 4. substances that attract natural enemies of a predator.
VOC interspecies communications that attract natural enemies of predators
There are many examples of this phenomenon. The 2011 publication A cry for help from leaf to root: Above ground insect feeding leads to the recruitment of rhizosphere microbes for plant self-protection against subsequent diverse attacks relates “Plants have evolved general and specific defense mechanisms to protect themselves from diverse enemies, including herbivores and pathogens. To maintain fitness in the presence of enemies, plant defense mechanisms are aimed at inducing systemic resistance: in response to the attack of pathogens or herbivores, plants initiate extensive changes in gene expression to activate “systemic acquired resistance” against pathogens and “indirect defense” against herbivores. Recent work revealed that leaf infestation by whiteflies, stimulated systemic defenses against both an airborne pathogen and a soil-borne pathogen, which was confirmed by the detection of the systemic expression of pathogenesis-related genes in response to salicylic acid and jasmonic acid-signaling pathway activation. Further investigation revealed that plants use self protection mechanisms against subsequent herbivore attacks by recruiting beneficial microorganisms called plant growth-promoting rhizobacteria/fungi, which are capable of reducing whitefly populations. Our results provide new evidence that plant-mediated aboveground to belowground communication and vice versa are more common than expected.”
An additional important point made here is that different predator and pathogen attacks can activate the same signaling pathway that upgrades complexes of defensive genes. The result is strengthened resistance to a variety of future attacks.
This is similar to hormesis as expressed on animals, a topic much-discussed in this blog. Going on with examples of VOC communications:
It has not been determined what plants are the intended target of the volatile signaling cues. It is possible that main targets are different branches of the same plant which are not sufficiently connected vascularly. However, the following several research papers show different ways plants can use volatiles to stimulate their own defenses against herbivores.
The case of hybrid poplar vs gypsy moth larvae shows how when one branch is attacked, other branches on the same plant respond by increasing their defenses
The 2007 publication Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores makes this point: “Plant volatiles play important roles in signaling between plants and insects, but their role in communication among plants remains controversial. Previous research on plant–plant communication has focused on interactions between neighbouring plants, largely overlooking the possibility that volatiles function as signals within plants. Here, we show that volatiles released by herbivore-wounded leaves of hybrid poplar (Populus deltoides × nigra) prime defences in adjacent leaves with little or no vascular connection to the wounded leaves. Undamaged leaves exposed to volatiles from wounded leaves on the same stem had elevated defensive responses to feeding by gypsy moth larvae (Lymantria dispar L.) compared with leaves that did not receive volatiles. Volatile signals may facilitate systemic responses to localized herbivory even when the transmission of internal signals is constrained by vascular connectivity. Self-signalling via volatiles is consistent with the short distances over which plant response to airborne cues has been observed to occur and has apparent benefits for emitting plants, suggesting that within-plant signalling may have equal or greater ecological significance than signalling between plants.”
Volatile signaling can work within the plant sagebrush, (Artemisia tridentata) and between plants up to 60 cm apart
The 2006 publication Damage-induced resistance in sagebrush: volatiles are key to intra- and interplant communication reports “”Airborne communication between individuals, called “eavesdropping” in this paper, can cause plants to become more resistant to herbivores when a neighbor has been experimentally clipped. The ecological relevance of this result has been in question, since individuals may be too far apart for this interaction to affect many plants in natural populations. We investigated induced resistance to herbivory in sagebrush, Artemisia tridentata, caused by experimental clipping of the focal plant and its neighbors. We found no evidence for systemic induced resistance when one branch was clipped and another branch on the same plant was assayed for naturally occurring damage. In this experiment, air contact and plant age were not controlled. Previous work indicated that sagebrush received less damage when a neighboring upwind plant within 15 cm had been experimentally clipped. Here we found that pairs of sagebrush plants that were up to 60 cm apart were influenced by experimental clipping of a neighbor. Furthermore, we observed that most individuals had conspecific neighbors that were much closer than 60 cm. Air contact was essential for communication; treatments that reduced airflow between neighboring individuals, either because of wind direction or bagging, prevented induced resistance. Airflow was also necessary for systemic induced resistance among branches within an individual. Reports from the literature indicated that sagebrush is highly sectorial, as are many desert shrubs. Branches within a sagebrush plant do not freely exchange material via vascular connections and apparently cannot rely on an internal signaling pathway for coordinating induction of resistance to herbivores. Instead, they may use external, volatile cues. This hypothesis provides a proximal explanation for why sagebrush does not demonstrate systemic induced resistance without directed airflow, and why airborne communication between branches induces resistance.”
When lima bean plants are attacked by herbivores, the plants give off volatiles thatattract natural predators of the herbivores, not only affecting the attacked plants but also the neighboring plants. But this does not happen when they are exposed to artificially wounded plants.
Lima beans vs. spider mites; mobilizing spider mites
The 2000 publication Herbivory-induced volatiles elicit defence genes in lima bean leaves is one of a number of relevant publications on plant VOC communications that started to occur in the early 2000s and that highlight the interspecies nature of the communications: “In response to herbivore damage, several plant species emit volatiles that attract natural predators of the attacking herbivores1, 2, 3, 4, 5. Using spider mites (Tetranychus urticae) and predatory mites (Phytoseiulus persimilis)1, 2, 3, 4, it has been shown that not only the attacked plant but also neighbouring plants are affected, becoming more attractive to predatory mites 3, 6 and less susceptible to spider mites 6. The mechanism involved in such interactions, however, remains elusive. Here we show that uninfested lima bean leaves activate five separate defence genes when exposed to volatiles from conspecific leaves infested with T. urticae, but not when exposed to volatiles from artificially wounded leaves. The expression pattern of these genes is similar to that produced by exposure to jasmonic acid. At least three terpenoids in the volatiles are responsible for this gene activation; they are released in response to herbivory but not artificial wounding. Expression of these genes requires calcium influx and protein phosphorylation/dephosphorylation.”
Lima beans vs. beetles; mobilizing predatory anthropods
A subsequent publication on lima bean plants appearing in 2007 was Within-plant signaling by volatiles leads to induction and priming of an indirect plant defense in nature: “Plants respond to herbivore attack with the release of volatile organic compounds (VOCs), which can attract predatory arthropods and/or repel herbivores and thus serve as a means of defense against herbivores. Such VOCs might also be perceived by neighboring plants to adjust their defensive phenotype according to the present risk of attack. We exposed lima bean plants at their natural growing site to volatiles of beetle-damaged conspecific shoots. This reduced herbivore damage and increased the growth rate of the exposed plants. To investigate whether VOCs also can serve in signaling processes within the same individual plant we focused on undamaged “receiver” leaves that were either exposed or not exposed to VOCs released by induced “emitter” leaves. Extrafloral nectar secretion by receiver leaves increased when they were exposed to VOCs of induced emitters of neighboring plants or of the same shoot, yet not when VOCs were removed from the system. Extrafloral nectar attracts predatory arthropods and represents an induced defense mechanism. The volatiles also primed extrafloral nectar secretion to show an augmented response to subsequent damage. Herbivore-induced VOCs elicit a defensive response in undamaged plants (or parts of plants) under natural conditions, and they function as external signal for within-plant communication, thus serving also a physiological role in the systemic response of a plant to local damage.”
When alders are attacked by the alder leaf beetle, their undamaged neighbor alders also respond with increased resistance to the alder beetle leaf herbivory. The lab experiments also supported field results. Defoliation of alders may trigger interplant resistance transfer, and therefore reduce herbivory in whole alder stands.
Alders vs. alder leaf beetles
Another 2000 publication that helped facilitate the swing back to acceptance of plant VOC communications was Defoliation of alders (Alnus glutinosa) affects herbivory by leaf beetles on undamaged neighbors: “The effects of defoliation of alder (Alnus glutinosa) on subsequent herbivory by alder leaf beetle (Agelastica alni) were studied in ten alder stands in northern Germany. At each site, one tree was manually defoliated (c.20% of total foliage) to simulate herbivory. Subsequent damage by A. alniwas assessed on ten alders at each site on six different dates from May to September 1994. After defoliation, herbivory by A. alni increased with distance from the defoliated tree. Laboratory experiments supported the field results. Not only leaf damage in the field, but also the extent of leaf consumption in laboratory feeding-preference tests and the number of eggs oviposited per leaf in another laboratory test were positively correlated with distance from the defoliated tree. Resistance was therefore induced not only in defoliated alders, but also in their undamaged neighbours. Consequently, defoliation of alders may trigger interplant resistance transfer, and therefore reduce herbivory in whole alder stands.”
Sound communications
Plants can also respond to vibrational communications. The 2014 publication Plants respond to leaf vibrations caused by insect herbivore chewing reports: “Plant germination and growth can be influenced by sound, but the ecological significance of these responses is unclear. We asked whether acoustic energy generated by the feeding of insect herbivores was detected by plants. We report that the vibrations caused by insect feeding can elicit chemical defenses. Arabidopsis thaliana (L.) rosettes pre-treated with the vibrations caused by caterpillar feeding had higher levels of glucosinolate and anthocyanin defenses when subsequently fed upon by Pieris rapae (L.) caterpillars than did untreated plants. The plants also discriminated between the vibrations caused by chewing and those caused by wind or insect song. Plants thus respond to herbivore-generated vibrations in a selective and ecologically meaningful way. A vibration signaling pathway would complement the known signaling pathways that rely on volatile, electrical, or phloem-borne signals. We suggest that vibration may represent a new long distance signaling mechanism in plant–insect interactions that contributes to systemic induction of chemical defenses.”
Continuing: “The effects of sound on plant growth and other traits have been recognized for decades, but the ecological significance of these responses is unclear. While plant responses to wind and touch have been examined and have clear adaptive significance (Chehab et al. 2009), plant responses to acoustic energy have largely been studied in the absence of an ecological context. For example, there is a long tradition of exposing plants to musical sound (Klein and Edsall 1965; Telewski 2006; Jeong et al. 2004). Although music influences growth and germination in some plants, music contains such a wide range of frequencies, amplitudes and fine-temporal patterns that its usefulness as an experimental stimulus is limited. More systematic studies have found that some frequencies have a greater influence than others (Telewski 2006). For example, young roots of corn grow towards the source of continuous tones, transmitted as airborne or waterborne sound, and respond optimally to frequencies of 200–300 Hz (Gagliano et al. 2012a). While these studies bring us a step closer to being able to link plant responses to acoustic energy to ecologically relevant sound sources, the experimental stimuli still remain far removed from those produced by natural sources of acoustic energy in the plant’s environment. — One of the most relevant sources of acoustic energy in the immediate environment of a plant is the rich community of plant-associated arthropods, including herbivores, predators, and parasitoids (Cocroft and Rodriguez 2005). Plant-borne vibrations provide a wealth of information about the activities of insects on plants. Within the abundant arthropod community on plants, many ecological and social interactions depend on the perception and production of plant-borne vibrations (Hill 2008). Some 200,000 species of insects communicate using substrate vibrations to locate mates, attract mutualists, or exploit plant resources (Cocroft and Rodriguez 2005). Many more insects and other arthropods use such vibrations to locate prey or avoid predators (Barth 1998; Castellanos and Barbosa 2006; Casas and Magal 2006; Virant-Doberlet et al.2011; Cocroft 2011). Chewing herbivores, in particular, produce characteristic, high-amplitude vibrations that travel rapidly to other parts of the plant. Predatory insects can use chewing vibrations to detect their prey from a considerable distance: for example, on soybean, the chewing vibrations of green clover worms elicited search by predatory stinkbugs 50 cm away (Pfannenstiel et al. 1995). We suggest that the vibrations produced by chewing herbivores are an important source of acoustic energy for plants. If plants can detect and use this conspicuous, reliable and rapidly transmitted source of information about herbivore feeding, tissues far from the site of attack could use feeding vibrations to respond quickly to the threat of herbivory. A vibration signaling pathway would complement the known signaling pathways that rely on phloem-borne signals, airborne volatiles, or electrical signals (Wu and Baldwin 2009; Mousavi et al. 2013). Here we test the hypothesis that plant responses to herbivory, in the form of induced chemical defenses, can be elicited by the mechanical vibrations produced by chewing caterpillars. We report that Arabidopsis thaliana plants exposed to chewing vibrations produced greater amounts of chemical defenses in response to subsequent herbivory, and that the plants distinguished chewing vibrations from other environmental vibrations.”
Root communications
This is a form of extended communications we humans do not have.
Touch and chemical communications can also occur via undergrownd root-connecting mycelial networks, e,g, networks of fungal filaments. The 2012 publication Mycorrhiza-induced resistance and priming of plant defenses reports: “Symbioses between plants and beneficial soil microorganisms like arbuscular-mycorrhizal fungi (AMF) are known to promote plant growth and help plants to cope with biotic and abiotic stresses. Profound physiological changes take place in the host plant upon root colonization by AMF affecting the interactions with a wide range of organisms below- and above-ground. Protective effects of the symbiosis against pathogens, pests, and parasitic plants have been described for many plant species, including agriculturally important crop varieties. Besides mechanisms such as improved plant nutrition and competition, experimental evidence supports a major role of plant defenses in the observed protection. During mycorrhiza establishment, modulation of plant defense responses occurs thus achieving a functional symbiosis. As a consequence of this modulation, a mild, but effective activation of the plant immune responses seems to occur, not only locally but also systemically. This activation leads to a primed state of the plant that allows a more efficient activation of defense mechanisms in response to attack by potential enemies. Here, we give an overview of the impact on interactions between mycorrhizal plants and pathogens, herbivores, and parasitic plants, and we summarize the current knowledge of the underlying mechanisms. We focus on the priming of jasmonate-regulated plant defense mechanisms that play a central role in the induction of resistance by arbuscular mycorrhizas.”
Linking into a mycelia network can have a profound positive effect on gene expression health state and disease resistance in the linked plants.
The 2007 publication Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots reports: “In natural ecosystems, the roots of many plants exist in association with arbuscular mycorrhizal (AM) fungi, and the resulting symbiosis has profound effects on the plant. The most frequently documented response is an increase in phosphorus nutrition; however, other effects have been noted, including increased resistance to abiotic and biotic stresses. Here we used a 16,000-feature oligonucleotide array and real-time quantitative RT-PCR to explore transcriptional changes triggered in Medicago truncatula roots and shoots as a result of AM symbiosis. By controlling the experimental conditions, phosphorus-related effects were minimized, and both local and systemic transcriptional responses to the AM fungus were revealed. The transcriptional response of the roots and shoots differed in both the magnitude of gene induction and the predicted functional categories of the mycorrhiza-regulated genes. In the roots, genes regulated in response to three different AM fungi were identified, and, through split-root experiments, an additional layer of regulation, in the colonized or non-colonized sections of the mycorrhizal root system, was uncovered. Transcript profiles of the shoots of mycorrhizal plants indicated the systemic induction of many genes predicted to be involved in stress or defense responses, and suggested that mycorrhizal plants might display enhanced disease resistance. Experimental evidence supports this prediction, and mycorrhizal M. truncatula plants showed increased resistance to a virulent bacterial pathogen, Xanthomonas campestris. Thus, the symbiosis is accompanied by a complex pattern of local and systemic changes in gene expression, including the induction of a functional defense response.”
Plants propagate electrical signals in response to artificial wounding. However, little is known about the electrophysiological responses of the phloem to wounding, and whether natural damaging stimuli induce propagating electrical signals in this tissue.
Here, we used living aphids and the direct current (DC) version of the electrical penetration graph (EPG) to detect changes in the membrane potential of Arabidopsis sieve elements (SEs) during caterpillar wounding.
Feeding wounds in the lamina induced fast depolarization waves in the affected leaf, rising to maximum amplitude (c. 60 mV) within 2s. Major damage to the midvein induced fast and slow depolarization waves in unwounded neighbor leaves, but only slow depolarization waves in non-neighbor leaves. The slow depolarization waves rose to maximum amplitude (c. 30 mV) within 14 s. Expression of a jasmonate-responsive gene was detected in leaves in which SEs displayed fast depolarization waves. No electrical signals were detected in SEs of unwounded neighbor leaves of plants with suppressed expression of GLR3.3 and GLR3.6.
EPG applied as a novel approach to plant electrophysiology allows cell-specific, robust, real-time monitoring of early electrophysiological responses in plant cells to damage, and is potentially applicable to a broad range of plant–herbivore interactions”
Multimodal communications involving mixed communications means between species both above and below ground
Now, we provide examples of interspecies plant communications of increasing complexity.
Underground mycelia networks can provide early warning between plants of an impending aphid attack.
Bean plants vs. aphids; mobilizing parasitoids using a fungal communications channel
The 2013 publication Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack reports: “The roots of most land plants are colonised by mycorrhizal fungi that provide mineral nutrients in exchange for carbon. Here, we show that mycorrhizal mycelia can also act as a conduit for signalling between plants, acting as an early warning system for herbivore attack. Insect herbivory causes systemic changes in the production of plant volatiles, particularly methyl salicylate, making bean plants, Vicia faba, repellent to aphids but attractive to aphid enemies such as parasitoids. We demonstrate that these effects can also occur in aphid-free plants but only when they are connected to aphid-infested plants via a common mycorrhizal mycelial network. This underground messaging system allows neighbouring plants to invoke herbivore defences before attack. Our findings demonstrate that common mycorrhizal mycelial networks can determine the outcome of multitrophic interactions by communicating information on herbivore attack between plants, thereby influencing the behaviour of both herbivores and their natural enemies.” We have at least four different species involved in this case.
Further complex plant communications and responses can lead to complex responses, such as a below ground insect attack leading to above ground protection against attack by insects of a different species.
The 2008 publication Plants as green phones: Novel insights into plant-mediated communication between below- and above-ground insects reports “Plants can act as vertical communication channels or ‘green phones’ linking soil-dwelling insects and insects in the aboveground ecosystem. When root-feeding insects attack a plant, the direct defense system of the shoot is activated, leading to an accumulation of phytotoxins in the leaves. The protection of the plant shoot elicited by root damage can impair the survival, growth and development of aboveground insect herbivores, thereby creating plant-based functional links between soil-dwelling insects and insects that develop in the aboveground ecosystem. The interactions between spatially separated insects below- and aboveground are not restricted to root and foliar plant-feeding insects, but can be extended to higher trophic levels such as insect parasitoids. Here we discuss some implications of plants acting as communication channels or ‘green phones’ between root and foliar-feeding insects and their parasitoids, focusing on recent findings that plants attacked by root-feeding insects are significantly less attractive for the parasitoids of foliar-feeding insects.”
Interaction communications among a plant, fungi, insects and other plants can be complex, multimodal and multidirectional
In the real world it gets a lot more complicated. The 2013 publication Two-way plant mediated interactions between root-associated microbes and insects: from ecology to mechanisms reports: “Plants are members of complex communities and function as a link between above- and below-ground organisms. Associations between plants and soil-borne microbes commonly occur and have often been found beneficial for plant fitness. Root-associated microbes may trigger physiological changes in the host plant that influence interactions between plants and aboveground insects at several trophic levels. Aboveground, plants are under continuous attack by insect herbivores and mount multiple responses that also have systemic effects on belowground microbes. Until recently, both ecological and mechanistic studies have mostly focused on exploring these below- and above-ground interactions using simplified systems involving both single microbe and herbivore species, which is far from the naturally occurring interactions. Increasing the complexity of the systems studied is required to increase our understanding of microbe–plant–insect interactions and to gain more benefit from the use of non-pathogenic microbes in agriculture. In this review, we explore how colonization by either single non-pathogenic microbe species or a community of such microbes belowground affects plant growth and defense and how this affects the interactions of plants with aboveground insects at different trophic levels. Moreover, we review how plant responses to foliar herbivory by insects belonging to different feeding guilds affect interactions of plants with non-pathogenic soil-borne microbes. The role of phytohormones in coordinating plant growth, plant defenses against foliar herbivores while simultaneously establishing associations with non-pathogenic soil microbes is discussed.”
There is a growing interest in all the complex interactions of plants with other plants and organisms and how this affects ecological systems.
The 2012 publication Root herbivore effects on aboveground multitrophic interactions: patterns, processes and mechanismsreports: “In terrestrial food webs, the study of multitrophic interactions traditionally has focused on organisms that share a common domain, mainly above ground. In the last two decades, it has become clear that to further understand multitrophic interactions, the barrier between the belowground and aboveground domains has to be crossed. Belowground organisms that are intimately associated with the roots of terrestrial plants can influence the levels of primary and secondary chemistry and biomass of aboveground plant parts. These changes, in turn, influence the growth, development, and survival of aboveground insect herbivores. The discovery that soil organisms, which are usually out of sight and out of mind, can affect plant-herbivore interactions aboveground raised the question if and how higher trophic level organisms, such as carnivores, could be influenced. At present, the study of above-belowground interactions is evolving from interactions between organisms directly associated with the plant roots and shoots (e.g., root feeders – plant – foliar herbivores) to interactions involving members of higher trophic levels (e.g., parasitoids), as well as non-herbivorous organisms (e.g., decomposers, symbiotic plant mutualists, and pollinators). This multitrophic approach linking above- and belowground food webs aims at addressing interactions between plants, herbivores, and carnivores in a more realistic community setting. The ultimate goal is to understand the ecology and evolution of species in communities and, ultimately how community interactions contribute to the functioning of terrestrial ecosystems. Here, we summarize studies on the effects of root feeders on aboveground insect herbivores and parasitoids and discuss if there are common trends. We discuss the mechanisms that have been reported to mediate these effects, from changes in concentrations of plant nutritional quality and secondary chemistry to defense signaling. Finally, we discuss how the traditional framework of fixed paired combinations of root- and shoot-related organisms feeding on a common plant can be transformed into a more dynamic and realistic framework that incorporates community variation in species, densities, space and time, in order to gain further insight in this exciting and rapidly developing field.”
Plants, predators and bacteria can form various alliances for survival. In some cases the plants come out ahead, in other cases they do not.
An example of the second kind of situation is described in a 2013 Science Daily article Microbes help beetles defeat plant defenses: “Some symbiotic bacteria living inside Colorado potato beetles can trick plants into reacting to a microbial attack rather than that of a chewing herbivore, according to a team of Penn State researchers who found that the beetles with bacteria were healthier and grew better. “For the last couple of decades, my lab has focused on induced defenses in plants,” said Gary W. Felton, professor and head of entomology. “We had some clues that oral secretions of beetles suppressed defenses, but no one had followed up on that research.” Seung Ho Chung, graduate student in entomology working with Felton, decided to investigate how plants identified chewers and how herbivores subverted the plants defenses. “I thought we could identify what was turning the anti-herbivore reaction off,” said Felton. “But it was a lot more difficult because we had not considered microbes.” According to Felton, the beetles do not have salivary glands and so they regurgitate oral secretions onto the leaves to begin digestion. These secretions contain gut bacteria. — Plant defenses against chewing insects follow a jasmonite-mediated pathway that induces protease inhibitors and polyphenol oxidase, which suppress digestion and growth. Plant defenses against pathogens follow a salicylic acid mediated pathway. When the antimicrobial response turns on, it interferes with the response to chewing, allowing the beetles to develop more normally.”
“Chung and Felton used tomato plants to identify exactly what was turning off the response to chewing. They note, however, that the Colorado potato beetle also attacks eggplant and potato plants. They report the results of their work in the current online edition of the Proceedings of the National Academy of Sciences. — The researchers allowed beetle larva to feast on antibiotic-treated leaves and natural leaves and found that on the antibiotic-treated leaves, the beetles suffered from the plant’s anti-herbivore defense, but on the natural leaves the larva gained more weight and thrived. Chung and Felton then investigated expression of genes in the anti-herbivore pathway and the production of enzymes. They found that the presence of bacteria decreases the anti-herbivore response. — The researchers also isolated and grew the bacteria from the Colorado potato beetle guts. They found 22 different types of bacteria, but only three types suppressed the anti-herbivore response. During a variety of other experiments, they found that in all cases presence of the bacteria that could suppress the anti-herbivore response led to healthier beetles.”
Chemical communication from a plant to members of another species can be designed to create an addiction which enforces a long-term symbiotic relationship between the plant and members of that species
Phony messaging is one dirty trick that can be used in nature. Another is creating an addiction. An example is that a plant can create an addiction that keeps members of another species supporting the plant. One such case is explained in a 2013 National Geographic article Trees Trap Ants Into Sweet Servitude“An evolutionary alliance between trees and the ants that guard them has a sinister explanation, a new study suggests, finding ants hooked on nectar. In Central America, ants act as bodyguards for acacia trees, defending them from weeds and hungry animals in exchange for room and board, one of the most iconic alliances in nature. (See “Ant Bodyguards Get Exclusive Contract From Trees.”) But Martin Heil of Cinvestav Unidad Irapuato in Mexico has found that the tree’s sugary snacks are laced with an enzyme that prevents the ants from eating other sources of sugar. One sip, and the insects are consigned to a life of indentured servitude. “It was surprising to me that the immobile, ‘passive’ plant can manipulate the seemingly much more active partner, the ant,” says Heil. The report illustrates how evolution keeps cooperative relationships among some species going, even when one partner is clearly reaping most of the benefits in the arrangement. (See “Video—Acacia Tree Ants.”)”
“Nectar Addicts:Heil compares the tree to a dairy company that sells lactose-free milk that has been chemically altered to render its customers unable to digest normal milk. Any customer who drank it would be forced to stick to that one lactose-free brand. Here’s how it works. Most of the sweet foods that ants eat, such as plant sap, are rich in a sugar called sucrose. The ants digest this with an enzyme called invertase, which breaks sucrose into smaller sugars. In 2005, Heil showed that all of the workers of the acacia ant Pseudomyrmex ferrugineus lack invertase activity and cannot digest normal sources of sucrose. Fortunately, the tree compensates for this impairment by secreting invertase into its nectar, providing the ants with a predigested meal. For this reason, the ants prefer acacia nectar over other sugar sources. But Heil suspected that this explanation was too neat. Why would the ants so heavily restrict their dietary options by losing an important enzyme? Weirder still, in 2008, Heil showed that the larvae have invertase, which becomes deactivated only when they turn into adults. Culprit Nabbed:Heil has now shown that the tree itself is responsible. Writing in the Ecology Letters journal, he reports that acacia nectar contains chitinase enzymes that completely block invertase. Shortly after the workers emerge from their pupae as adults, they take their first sip of nectar and their invertase becomes irreversibly disabled. “Ain’t nature grand?” says Todd Palmer of the University of Florida, who studies ants and acacias. “What looks from the outside as another case of digestive specialization appears to be a sneaky manipulation on the part of the acacia to increase ant dependence.”
A given species of plants can adopt multiple inter and intra-species communications defenses against various predators and incorporate these into multi-aspect overall defensive strategies.
Above, we describe how Acacia trees chemically enlist ants into servitude to help themselves. This is part of a multipronged defense strategy used in Africa’s savannas and woodlands by Acacia tortilis against being eaten by Giraffes. The story is told in NO PLACE TO RUN NOT PLACE TO HIDE from GardenSmart.
In summary, Giraffes love gobbling Acacia leaves, and can consume 140 pounds of leaves a day. They can run 35 miles per hour and find the leaves very nourishing. The tees can’t run away or hide from giraffes but have developed these defenses: 1. A system of tough 4-inch long straight and curved thorns, 2. The ants mentioned in the previous item “A second line of defense is not readily apparent until contact is made with a protected plant. Ants – biting, stinging, swarming, and ready to give their lives in defense of the colony and turn a mouthful of foliage into a painful experience. Many plants draw ants into their canopies with the lure of nectar found in nectaries on the leaves. But certain species of acacia such as the whistling thorn (Acacia drepanolobium) go one step further by supplying room as well as board in the form of swollen thorn bases. The ants hollow out the thorns while green and become nest sites and living quarters, domatia. The colony readily defends its home tree against any and all intruders be they mammal, insect or plant. Pheromone scent trails rally the troops to battle and may serve as a warning to nearby browsers that these leafy morsels are served with a bite.” 3. A third line of defense, this one being again chemical involving tannins. “Tannins inhibit digestion by interfering with protein and digestive enzymes and binding to consumed plant proteins making them more difficult to digest. Herbivores have various strategies for dealing with tannins, some more successful than others.” 4. The plant may produce other toxic substances- “Under certain conditions, acacias build up levels of a toxin, known as Prussic acid or hydrocyanic acid or hydrogen cyanide (HCN). — Ruminant animals are more susceptible due to certain enzymes found in their digestive tract. This potent poison interferes with oxygen use at the cellular level causing death by asphyxiation.” 5. A fifth line of defense is warning other trees through release of ethelene gas. “Upon tearing away at the protein-rich foliage, the torn leaf surfaces emit the gaseous hormone ethylene, alerting other plants within 50 yards to increase tannin production in order to thwart the foraging mega-herbivore. Sensing the menu change the giraffe moves upwind dining on plants that have failed to catch the drift. More than the adjoining vegetation may detect this call to arms. Passing carnivores pick up the invitation, resulting in a hasty retreat or, at the very least, a change of plans for the browser. 6. Finally, a sixth level of defense is internal signaling in the acacia plant which fosters rapid regrowth. “Often, branches that have been pruned and stripped of leaves grow back more vigorously than unbrowsed branches. Tipped branches send out many side shoots, each with new leaves that seasonally provide even more nutritious browsing than before, while increased leaf surface allows increased energy production. Repeatedly done this would be analogous to a gardener shearing a hedge. This mutuality outcome works out for both parties, over the short term anyways. Browsed stems are more likely than unbrowsed ones to be dead the next year and repeated heavy tissue loss and replacement could ultimately deplete the trees available resources unless some compensatory source of nutrients becomes available.” Giraffes have developed counter-measures to some of these defenses. So they and arcacias live in a dynamic ecological balance. As we do too, though we might not like to admit it.
Increasing healthful phytosubstances in fruits and vegetables via plant signaling approaches
Utilization of knowledge of plant pest-response signaling could facilitate improved planting and cultivation methods for organic farming. For example, improved ways might be developed for planting crops so they will signal to each other with VOCs if there is a pest threat. Such might do a better job than chemical pesticides. A just-published meta-study appearing in the British Journal of Nutrition indicates that the nutritional and health benefits of doing this could be very great(ref). The report identifies significant differences between organically grown and conventionally grown fruits, vegetables and grains. organically grown fruits, vegetables and grains are likely to have 18 to 69 percent higher concentrations higher concentrations of beneficial phytochemicals and are much less likely to contain toxic pesticides. According to a Washington State University press release: “The study looked at an unprecedented 343 peer-reviewed publications comparing the nutritional quality and safety of organic and conventional plant-based foods, including fruits, vegetables, and grains. — Most of the publications covered in the study looked at crops grown in the same area, on similar soils. This approach reduces other possible sources of variation in nutritional and safety parameters. — In general, the team found that organic crops have several nutritional benefits that stem from the way the crops are produced. A plant on a conventionally managed field will typically have access to high levels of synthetic nitrogen, and will marshal the extra resources into producing sugars and starches. As a result, the harvested portion of the plant will often contain lower concentrations of other nutrients, including health-promoting antioxidants. — Without the synthetic chemical pesticides applied on conventional crops, organic plants also tend to produce more phenols and polyphenols to defend against pest attacks and related injuries. In people, phenols and polyphenols can help prevent diseases triggered or promoted by oxidative-damage like coronary heart disease, stroke and certain cancers.” (We note that “antioxidants” here actually refer to plant polyphenols that can upgrade human antioxidant-response genes.) “Overall, organic crops had 18 to 69 percent higher concentrations of antioxidant compounds. The team concludes that consumers who switch to organic fruit, vegetables, and cereals would get 20 to 40 percent more antioxidants. That’s the equivalent of about two extra portions of fruit and vegetables a day, with no increase in caloric intake. The researchers also found pesticide residues were three to four times more likely in conventional foods than organic ones, as organic farmers are not allowed to apply toxic, synthetic pesticides. While crops harvested from organically managed fields sometimes contain pesticide residues, the levels are usually 10-fold to 100-fold lower in organic food, compared to the corresponding, conventionally grown food.”
Over the years we have written many articles in this blog about the health-producing properties of plant producing phytosubstances and how they activate beneficial xenohormetic pathways in humans via the Nrf2 pathway, and affects on Histone acetylation/deacetylation etc.(ref)(ref) (ref). Also, we have written about how stress signaling can be used to induce harvested fruits and vegetables to enhance their content of such phytochemicals, our articles on the xenohormetic live food hypothesis(ref)(ref)(ref).
Wrapping it up
We have been selective in picking the information to be presented in this blog, our intention being to offer a general introduction to plant communications with emphasis on communications potentially involving other plants or members of other species. We see plant communications as relevant to the intention of our team members to help create a Grand Unified Theory of biology and aging. We will probably follow this with other blog entries related to specialized topics related to plants such as the molecular biology of plant communications. Even more exciting, we might explore what is known about how aging works in plants. The longest-living species by a very wide margin are plants. Giant sequoia trees are reputed to live 3,200 years and creosole bush 11,700 years(ref). BristleCone Pine trees can live for up to 5.000 years. For a little more on them, you can see Vince’s 2013 PowerPoint presentation The Prospects that Emerging Science Offers Us for Longer Healthy Lifespans which you can download by clicking Newsciencesaging.
Can we expect to live longer and longer as the first part of this century rolls by? I think so, probably by a large amount. We about doubled our human lifespans in the course of the last 100 years. We can probably do this again, this time in as a little as 50 years for society as a whole, perhaps 30 years for those who systematically pursue this as an objective. This blog entry is about how that process can be systematized and speeded up, for individuals who wish to live very long healthy lives and for society as a whole. It is not about the many specific knowledge based interventions and changes that will lead to longer lives – matters that we have sought to cover in other entries in this blog.
Will this extension of human lives be because of basic new scientific breakthroughs? Yes, but only in part. Collectively, such breakthroughs are likely to be important. Curiously though, I don’t think that any single such breakthrough will make an immense difference. What will matter is a process issue, and whether and how such breakthroughs are applied is only one consideration.
So, how then will it happen? I argue here that extended longevity is likely to happen via a number of incremental steps,probably small ones at that. Most will involve improvements in lifestyle and diet. Others will involve selective application of stresses and consumption of health-producing phytosubstances and selected dietary supplements.
Can you or I as individuals accelerate this process? I believe the answer is generally yes. To the extent that the steps are science and knowledge-based and can be systematically identified and socially pursued, the process will be accelerated.
How? I think you can move along the increasing longevity curve by pursuing a long string of incremental lifestyle and dietary modifications over time, each of which may seem to produce only modest results. Some steps may seem to be very tiny and insignificant, such as getting up from the computer and walking around a bit every hour.
Does the knowledge to do this exist now? Yes, again I think so. You can distill much of it from our blog entries. And more is constantly more being learned as time rolls on.
I don’t get it. What exactly do I need to do?
Here is a story of amazing life extension that suggests the answer to this question.
Let’s start by talking about, my grandmother’s 1950 Chevy Bel Air, purchased new. This Chevy was a neat looking and zippy little car. I learned to drive on it and loved it dearly. However, like other cars of that era it tended to rust out and develop progressively serious problems starting in about a year. Like the other cars I will be talking about, it was purchased new. If you were middle class and lived in Detroit you were expected to turn your car in every year for a new car, or at least every 2-3 years. Most cars did not survive the junk heap for more than 4-5 years. A three-year-old car was a seriously old car and you could expect to put less than 50,000 miles on it before it died. The Bel Air got 13 -17 miles to the gallon of gas. My wife had a five-year-old Chevy when I was in my 20s. We had to get rid of it because the floor of the backseat had rusted out to the point where a small child could fall through it down to the road.
Image may be NSFW. Clik here to view.1959 Studebaker Lark, this picture like my first new car. Actual lifespan 3.4 years. RIP 1961 with 28,900 miles on it. Died of multiple complications in the engine, transmission and the frame coming apart. Back then, you junked a car when it was clearly not economical to repair it. Image source. It was sure a neat looker.
The problem of quality and short lifespans was not one just of US made cars; it was worldwide and applied to most cars in every price range. My shiny new Triumph Herald Convertible coupe – a sporty car made in England – purchased in 1968 was junk and had to be scrapped in 1972 with only 34,000 miles on it. It was built in a tradition of English craftsmanship and had a beautifully polished mahogany dashboard panel. Now a classic car, the Beatles had one. However, underneath the sporty exterior was a vintage mechanical system with design probably going back to the 1930s and 1940s. The engine was remarkably fussy and always going out of tune and the gears were traditional square cut which meant that you had to carefully push the clutch in twice every time you wished to shift gears.
Image may be NSFW. Clik here to view.The car in the picture is like our 2012 Subaru Impreza which we purchased new in 2011, an existing family car. We expect it to have a lifespan of 15-20 years. Image source We have had this car for almost three years now, but it is still a very young car. It will probably go a 3-4 more years before it develops any significant issues. We expect to be able to clock up between 250,000 and 300,000 miles on it. It gets 35 miles to the gallon on the highway, 28 city driving.
The car in the picture is like our 2005 Subaru Impreza which we purchased in 2004. Expected lifespan: 15-20 years, perhaps 200,000 miles Image source This 10 year old car is still healthy, vigorous and a reliable family workhorse with no known problems at 100,000 miles. No sign of rust. If it were human it would be like a healthy 35-year-old.
The big breakthrough
What was the big scientific or technical breakthrough that made the difference in lifespan and performance between the earlier cars and our Subarus? Lifespan extension of a factor of at least four and MPG improvement by a factor of two? None! In fact, it wasn’t any single big scientific or engineering breakthrough. The difference is because of thousands of incremental improvements made year after year in just about every component and system. Virtually everything has been improved to make cars more reliable, last longer and operate more economically. Different lighter more rustproof metals are used, and the fenders and bumpers are made of tough light plastic that can never rust. Exposed metal surfaces are treated with sophisticated chemicals to prevent rust. The metals in the engine are different as well as the electronic ignition and gas vaporization systems, and dimensional tolerances are much more precise in the newer cars. The gasoline and fluids used in the newer car are different, the automobiles are much safer on multiple dimensions. In 1950 and 1968 they did not have seatbelts, airbags, antilock brakes, reinforced chassis, let alone radar, GPS, computer-based engine, all-wheel drive, catalytic converter, safety monitoring systems, etc. And the maintenance requirements for the earlier cars were much greater than for our current cars. The newer cars do not require distributor or carburetor cleaning or adjustments because these no longer exist.
The health and longevity of the newer car was the result of a breakthrough in fact, but a breakthrough of a different kind. That breakthrough is known as Total Quality Management (TQM), a systematic approach to both design and manufacturing that seeks to achieve constant incremental improvements based on measurements, analysis and better integration of design and manufacturing processes. This approach was invented in the US but adapted early on by the Japanese automobile industry. What happened some six or more decades back is that Toyota Motors decided to break ranks with tradition and make better cars cheaper. This was the only way they could compete in the world market and in the US. By fifty years ago, Toyota motors was already heavily into quality control. It had adapted a TMQ organizational lifestyle and had implemented a Creative Idea Suggestion System. It had a corporate philosophy of customer first and quality first. See this article. US companies had the big advantages of cheap available raw materials and less shipping costs. The Japanese companies had the advantage of being smarter and adapted TMQ.
The US automobile industry centered in Detroit had a traditional philosophy of low-cost manufacturing, emphasis on style and marketing, and was happy with the idea that people had to trade in their cars after a year or two. Growing up in a middle-class family in Detroit, I knew from a very young age that trading in your car every year two was absolutely the expected thing to do in the 40s through the 70s. In the US automobile industry, it was known for sure that quality could come only at greater cost. So, competitors could not create it and have longer-lasting cars without those cars costing significantly more. Not to worry. This turned out to be wrong. TQM was based on the idea that it is possible to redesign automotive components for both quality and easier manufacturing and that far better automobiles could be made with less cost. Of course you had to keep working on that year-after-year, and that is what the Japanese companies did. In the 1980s and through the 1990s the US automobile industry was losing more and more domestic market share to the Japanese, and the US cars were achieving the reputation of being relatively junky.
As time progressed, other Japanese auto companies and eventually the US automobile companies were forced to adopt more and more elements of TQM and its successor quality philosophies in order to remain competitive in the world market. So, our homegrown automobiles also last a lot longer now than they used to. In the process the US companies permanently lost a great deal of market share for autos and small trucks to Japanese, European and other Asian companies.
Quadrupling human lifespan, or just doubling it?
Some 62 years elapsed between 1950 and 2012. Now, suppose we seriously applied something like TMQ to human health and longevity for the next 62 years. Do I think we could quadruple human healthspan and lifespan, like we did for automobiles? The idea seems completely preposterous. However, we might simply go for doubling our expected lifespans. Still preposterous? Perhaps not. And for a growing cohort of health-aware people. I would go for accomplishing this in 30 years, not 62.
I lay out how some basic TQM principles might be applied to health and longevity below. First, however, I would like to cast doubt on the idea that any single scientific breakthrough could by itself make a major difference in human healthspan or lifespan – much as I would personally love to see that happen.
Why individual scientific breakthroughs are unlikely by themselves to make major differences in human healthspans and lifespans.
I argue this point from two different systems perspectives.
The systems nature of human biology. As a corollary, the nature of human homeostatic feedback systems.
In the case of automobiles many different approaches across many systems had to be employed to extend their life spans. No single intervention could begin to do the trick and no single intervention made much of the amazing entire difference. And with every major shift in design and manufacturing process new problems and new opportunities for further improvement surfaced. Biologically speaking, we are vastly more complex systems subject to many more constraints and system interdependencies. There is a large multiplicity of factors involved in maintaining health and aging. Some of these need to be addressed independently, some considering their interrelationships.
Homeostatic feedback systems in the human body constantly seek to restore a large number of working balances such as related to cell chemistry, electrolyte balance, protein expression, etc. I speculate, therefore, that any intervention which would have a major life extending capability would activate one or more restorative feedback loops which would mitigate against the impact of that intervention being too great. At least, this seems to be true historically. As far as we know, life-extending interventions which may more than triple lifespans of simple organisms like nematodes can add only 5% to 15% to lifespans of complex organisms like us. It has been said that if we could wipe out the three major disease killers of old people – heart diseases, cancer and dementias, that might add only 6 or 7 years to our expected lifespans. That is because of the multifactorial nature of aging.
In generating blog entries I have often pointed out that pursuing multiple small interventions may be more effective in generating health and longevity than seeking any single blockbuster approach. For example, in the 2011 blog entry Age-related cognitive decline: focus on interventionsI said in summary “First, there is not one simple approach to promoting memory and mental acuity, neurogenesis and healthy nerve cell metabolism, nor is there even a small handful. To get the best results it is likely that a large number of approaches should be pursued synergistically.” I believe this applies across the board to the other issues of aging such as metabolic health, averting cancers, frailty, sarcopenia, DNA degradation, etc.
The systems nature of human societies. As a corollary social homeostatic feedback systems
For the automobile, TQM involved evolution in automobile design, manufacturing, supplier enterprises, transportation systems, and customer expectations. Profound changes were required not only in systems and procedures but in every organization involved in the supply chain and especially in mindsets. These changes involved such factors as employee participation, effective use of human resources and the creation of many information feedback circuits. Our health and longevity is to a major extent driven by social systems, norms, expectations and habits. The most important contributions to public health and longevity historically have been societal ones, such as clean water, and sewage systems, cleaner air, cessation of smoking, inoculation and better sanitary habits. Gradual raising of awareness and implementing new initiatives tends to take a very long time – decades or even centuries. We may be able to speed this process up some when it comes to health, but we are likely still to be looking at decades
Moreover, any positive measure that threatens established patterns and vested interests will be fiercely resisted and slowed. We have seen this, for example, and the tobacco industries’ advertising and lobbying initiatives against the cessation of smoking – a battle that is still ongoing after more than 60 years. We see this kind of opposition to the idea of global warming and initiatives to switch to renewable energy sources. Similarly, the current public health initiatives to bring diets in the US into a better balance to limit the epidemic of diabetes and its metabolic cousins has been confronting fierce resistance from the corn sugar industrial lobby, fast food suppliers, and producers of industrial food products. The resistance is seen in our agricultural subsidy system, in our laws, in our legislatures, in our courts, in massive advertising campaigns, and in the minds of many influential people
Resistance to change is a characteristic of human cultures, no matter whether it comes from the Pharaoh, from the Pope, from the cigarette industry or from the corn sugar lobby. It makes no sense to bemoan that fact, but it can be helpful to recognize it. What I am suggesting is that if a perfect lifestyle and diet for longevity could be identified today (and I think we may be getting close to being able to do that), getting most people to buy into that lifestyle and diet might take another 50 to 100 years. So it may take a very long time for any given shift and health-related patterns to take hold in the general society. The good news, however, is that individuals can take health matters into their own hands and that multiple little improvements can piled on one another moving us right along the greater longevity curve..
The key point is that although there is a remarkable difference in life expectancy between my grandmother’s Chevy or my Triumph Herald and our current Subarus this came about through numerous small improvements over the years, each one of which could be dismissed as relatively unimportant. These all took place within a powerful context of relentless improvement which has been called TMQ. We can create such a context for health and longevity.
Some of the reasons why this process worked so well for automobiles are also relevant to how we can create constant improvements in our own health and longevity:
The knowledge and technology to make the necessary quality improvements to get where we are now with cars simply did not exist in 1950, or were unfamiliar to the industry and prohibitively expensive. For most people today, radically extending their health and lifespans looks that way
The automobile industry had an enormous investment in supply and manufacturing capabilities in 1950. They could not afford to scrap all these in one immense step, even if they knew how to make the necessary improvements which they did not. The same can be said for today’s massive healthcare system.
The marketplace in 1950 expected automobiles to be cheap, to be shiny and fashionable when they were new and to wear out or deteriorate rapidly. Today, we venerate youth and still expect sickness and frailty to start at age 70 or earlier.
The cost of gasoline was so low that mileage of an automobile was not an important cost factor. We did not see the environmental costs of global warming. Today the cost of industrial-produced food is low and we mostly do not see the associated health and longevity cost.
What was all-important to transform the situation was to be on a path of constant improvement – TQM. The same can hod for personal health and longevity
The yearly improvements in the automobile industry were affordable, and dictated by a changing competitive environment. Many of us are already highly competitive when it comes to one health dimension – fitness. We might broaden the definition of that to go beyond exercise.
New technologies that contribute to product quality could be embodied in the auto manufacturing process as they became available. These included computer aided design, new metals and composite materials, computer-aided manufacturing, robotics, and innovations in management such as just in time production. In this blog, we report on a stream of important discoveries and innovations in health and longevity as they become available. We also strive to flag those which we believe can be embodied in a personal health and longevity regimen.
Is it possible to employ an approach like TQM to enhance human health and longevity?
At first look it could be argued that the answer is no, laudable though the objective may be. We can’t do what the automobile industry did. That is we cannot redesign human beings component-by-component for health and longevity. And we can’t redesign the human birth process that makes us. We have to accept ourselves as we are, the products of millions of years of evolution. Most of our genes are the ones we started out with when our condos were caves.
This is not to be the end of the story, however. While our genes are mostly fixed, our epigenome is not and can be affected heritably by our interventions. We can definitely affect the directions of our evolution and probably the rate of positive evolutionary changes by epigenetic interventions. It is now known that epigenetic interventions can serve to promote or inhibit gene mutations. Aging involves a lifelong pattern of changes in DNA methylation, culminating in hyper methylation of GPC island promoter sites, and otherwise global gene hypomethylation. But this methylation too can be profoundly affected by epigenetic interventions. In various blog entries we have discussed how this might take place through utilizing histone deacetylase and histone acetylase phyto and other substances. For example, check out our three-part series on the epigenetic’s of cancer and aging and how those two deadly dragons can be seriously slowed Slaying Two Dragons with One Stone – How to Prevent Cancer and Aging with the Same Strategy – PART 1, PART 2, and PART 3. And, as I write this my colleague Jim Watson and I are planning new blog entries on epigenomic interventions.
On the one hand, our basic human design might impose restraints on our longevity that we cannot remove. An example could be lifelong accumulation of DNA damage and non-functional or age-related dysfunctional modifications to our DNA like transpositions and ALU repeats and circular DNAs and proliferating dysfunctional RNA species. On the other hand, we have extensive biological machinery for the repair of DNA and control of our DNA. We are beginning to understand interventions that may keep this machinery powerfully activated as we reach advanced ages. Every week brings new discoveries.
In summary, we do not know if there are basic design constraints that ultimately limit our lifespans and, if so, the ages at which these constraints become active. And, we do not know whether there are constraints that we can’t get around. Given that our average lifespans have multiplied by a factor of four from early prehistorical periods to today we may be able to keep living longer and longer lives for some time to come. We will probably only know our real lifespan barriers as we come to them.
A TQM approach to health and longevity
I am suggesting that individuals can pursue a philosophy similar to TQM for enhancing their personal health and longevity. TQM is basically an organizational strategy but it embodies critical elements that also can be adapted by an individual. These include:
A deep contextual commitment torelentless improvement in health and longevity. Such intentionality to support quality did not really exist in the automobile industry prior to TQM. Most people today either have no such intentionality today with regard to health or would know how to implement it if they did.
There have been multiple initiatives to integrate TQM into the processes of healthcare organizations and institutions (ref)(ref). These include public health organizations(ref), laboratories(ref), HMOs(ref) and hospitals(ref).
However, my focus in this blog entry is on a relatively new concept – how an individual in the context of his or her own family and network associations can apply the principles of TMQ to his or her own personal health and longevity.
A commitment to seeking and applying knowledge and the best available technologies to achieve steps of improvement. This knowledge spans the range from the latest scientific understandings relating to biology and health on the one hand to specific information about an individual’s health conditions on the other hand. The commitment extends also to going ahead and experimentally doing things that might enhance health, and seeing if they work. The process has inherent uncertainty and risk. Personally, I find it very exciting.
Use of strategy, data, and effective communications to integrate the quality discipline into the culture and activities of the individual or organization concerned.
For an individual concerned with personal longevity this involves developing a strategy which might include learning about specific personal disease processes, instituting changes in lifestyle and dietary patterns, learning more about longevity interventions, etc. It can include steps to obtain ongoing data as to how well that individual is doing given his strategic path. This could involve use of medical tests such as inflammatory indices, and data related to daily exercise such as can be provided by a FitBit. It can involve increasing use of cellphone health and fitness apps. An individual needs to communicate with members of his or her family and other supporting people and institutions like healthcare personnel to assure that his or her intentions and activities are understood and supported.
4. A focus on the customers’ needs and wants, rather than those of a supplier or sponsor.
For an individual seeking a longer healthier life, the customer and the major supplier are the same. Any conflict between supplier and customer needs can and must be resolved on a personal basis. Being a busy person, as a supplier I like the idea of eating fast and easy-to-grab meals. As a customer, I want to consistently take care of my health. As a supplier for a family birthday party, I am expected to provide a rich cake and ice cream. As a customer, I see it important for me and my family members not to eat these and instead eat more fresh fruits and vegetables.
For supplier institutions there may be conflicts between focusing on their own wants and needs and those of their customers or other intermediaries. Pharmaceutical companies need to maximize their profits, and this happens best when there are drugs that are not disease cures but are required lifelong for maintenance of health in the continuing presence of the disease. Seriously preventing any age-related disease like diabetes would hit the bottom lines of some pharma companies horribly. As mentioned, companies that make or serve unhealthy food can be expected to defend the status quo vigorously by lobbying, legal actions and advertising. Similarly to what the US automobile industry did in the 1950s and 1960s, and what the tobacco industry did for many decades, they can be expected to rely on massive advertising to maintain their marketing positions and obfuscate health considerations. As pointed out above, it may take many decades to resolve such conflicts in the customer’s interests.
5. Process-centered.
A fundamental part of TQM is a focus on process thinking. A process is a series of steps that take inputs from suppliers (internal or external) and transforms them into outputs that are delivered to customers (again, either internal or external). The steps required to carry out the process are defined, and performance measures are continuously monitored in order to detect unexpected variation.
For an individual, being processed centered implies focusing on every activity and habitual pattern that relates to health and seeing if it can be improved. In my personal case it is useful that my office is on the upper floor of the house is requiring me to climb on the average of a dozen flights of stairs every day. Everything related to movement, posture, patterns of activity, eating and social relationships is up to for grabs. That is, I ask myself if there is a variant of that activity that I know from science may be better for me. For example, does my pattern of research and writing on a computer support me getting up and moving around every hour? Does my daily rhythm allow for at least 45 minutes of vigorous exercise every day combined with other physical challenges throughout the day? Would I be better off shifting my treadmill exercise from the early evening to the morning? I may make a half-dozen shifts every week and see how they work for me, making some permanent and dropping others. And I think about others, e.g. running up the hill every day to get the mail, something I’ve been leaving for my wife to do. It could be doing a few stretching exercises every morning. And there are a huge number of process steps connected with shopping for foods and eating. A recent shift was from using regular milk to almond milk and from consuming sugar-rich raisin bran cereal to consuming oatmeal. Also I have become increasingly aware of stresses or absences of the same, recognizing the importance of experiencing stresses for health and vitality. I may deliberately choose to be rather uncomfortably cold or hot as part of my daily rhythm for this purpose.
Just like there was a myth that increasing quality in an automobile could only happen at greater cost, there is a myth that paying conscious attention to health can come at a cost of quality of life. I have found this to be completely untrue. While I thoroughly enjoyed breakfasting on cold milk and crispy raisin bran, I enjoy the warm oatmeal and almond milk with blueberries and walnuts even more.
6. Persistent confrontation and overcoming of barriers
As mentioned above, social, personal, and organizational feedback mechanisms operate powerfully in the background to maintain the status quo. In the two decades or more that efforts have been made to implement TMQ in healthcare organizations, for example, multiple barriers are still being encountered and have to be overcome. See the 1991 publication Organizational barriers to quality improvement in medical and health care organizations. “The barriers are categorized as technical, structural, psychosocial, managerial, and goals and values. Following a mapping of the barriers, education, training, and research and development needs to support quality improvement are identified.” Much has been written since 1965 about improving the quality of healthcare delivery and overcoming the multiple existing barriers – though there is still a very long way to go. See this list.
As to barriers to radical life extension, you might enjoy reading a story that I invented in the blog entry Getting the world ready for radical life extension. The Story of the X-Pill starts out: “A team of distinguished university researchers uses molecular engineering to create a substance X that appears to activate an evolutionary genetic pathway affecting the expression of hundreds of aging-related genes. When tested, the substance doubles the life of laboratory mice and rats. Based on solid theoretical considerations, it appears that X could be the basis for making an anti-aging pill for humans. Extrapolating up from the mice and rats, it appears in theory that people who start taking the X pill on a daily basis in their 40s will double their average life spans. They will, the reasoning goes, continue to age but at a much slower rate. They will still get the diseases and problems of old age but, on the average, much later. At age 110 they should be about as healthy as people are today at 55. — So, the researchers raised venture capital to start up a biotech company to manufacture X pills and sell them to the public.” What happened in the story? In short, proponents of the X pill confronted so many barriers and challenged so many vested interests that eventually the situation for them and their company was disastrous. The public never got to reap the benefits of the X pill or to see if it actually extended lives. Read the story for the details.
For an individual intending to extend his or her personal healthspan and lifespan, barriers can include 1. Lack of support of family members, and others and social isolation, 2. Opposition to what they are doing by officially sanctioned healthcare providers, 3, Lifelong personal habits and patterns that remain unexamined, 4. Confusion or uncertainty as to what steps could or could not personally be helpful, often made worse by conflicting advice of experts, 5. A perceived lack of time to do the necessary things such as exercise, 6. Lack of the infrastructure support, such as available paths or sidewalks for walking or lack of convenient suppliers of fresh fruits and vegetables, 7. Skepticism and ridicule by others, 8. Lack of a sufficient knowledge base about human biology, 8. Absence of easy to interpret measurements of health indicators, such as inflammatory state, overall REDOX state, degree of activation of certain critical proteins such as NF-kappaB, IGF-1, NRF2, etc. (It is interesting that much current efforts is being addressed to overcoming this barrier, including Moonshot, a Google initiative that is searching for a reliable set of biomarkers for general health and aging(ref).) And 9. absence of knowledgeable and supportive people the individual can communicate with. Part of the reason why creating health is an incremental process is that such barriers have to be confronted one at a time, and may re-arise in different contexts.
7. Integrated system
For organizations: “Although an organization may consist of many different functional specialties often organized into vertically structured departments, it is the horizontal processes interconnecting these functions that are the focus of TQM. Micro-processes add up to larger processes, and all processes aggregate into the business processes required for defining and implementing strategy(ref).”
Related to the biology of health and aging: as we have often pointed out there is no center to biological organisms or to aging. All systems are interconnected and considering any health intervention requires taking into account the impact of that intervention on all systems. In some cases there can be unforeseen and unwanted impacts and therefore systematic monitoring and evaluation of any introduced changes is necessary.
Regarding individuals seeking to improve their health and longevity, it is important that individual recognizes that here she is a participation in integrated social structures that can either aid or serve to resist moving forward with such improvements.
8. Empowerment
For organizations, it has been learned that empowerment of all employees to maximally contribute to the organization’s objectives is very important for successfully achieving TQM. Employees and all other individuals participating in the organization must be viewed not as cogs in a predefined machine, but rather as contributing components of a dynamic constantly self-adjusting organism. This requires empowering individuals to contribute to the maximum extent they can.
For an individual seeking to improve his or her health and longevity, the same applies to all important members of his human network, be they family members, close friends or colleagues, professional associates or healthcare providers. This means a requirement for sharing of personal health and longevity goals and objectives, and getting critical people in a personal support network to buy into supporting them. To be effective, the empowerment must be mutual.
9. Supply chain management
An important aspect of TQM in manufacturing organizations is paying careful attention to the supply chain – providers of raw materials and components, shipping organizations, wholesalers and retailers, etc. It is important to make sure there is alignment in plans and strategies and well articulated communications to support that alignment. An innovation in this regard has been just in time manufacturing, a process by which components used in manufacturing, say automobile fenders, arrive at an assembly plant just in time for them to be used, rather than stockpiled before hand. This facilitates major cost reduction connected with inventory control, more flexible manufacturing, and reduced headaches associated with inventory storage and management. Even better, using the computer communications it can be possible to assure that the fenders that arrive at an assembly plant on a given day match the models and colors of automobiles that will be assembled that day.
A biological analogue of this situation applies to eating and health. A major cause of obesity is the stockpiling of calories which are converted into fat as a result of eating which is not synchronized with actual metabolism. Obesity can thus be regarded as a problem of dietary inventory control – lack of dietary supply chain synchronization with the metabolic operation of the body. The healthiest situation involves careful articulation of dietary input, both with respect to the specific components consumed and the amounts, to match actual daily metabolic expenditures.
10. Innovation and pragmatic responsiveness
There is no such thing as a general agenda for improvement according to TQM. Every organization exists in its own ecology of other organizations and the steps to forward its TQM objectives at any given time will be unique to it. Changes in environment, customer expectations, or the availabilities of new technologies will pragmatically trigger steps of improvement. So, innovation in processes is the rule rather than the exception and is customized to the particular internal and external situation of a particular organization at a particular time.
Pretty much the same holds for individuals seeking to further their health and longevity. There is no general rule as to sequence of steps which need to be pursued but rather what is important is the ability of the individual to discern what the problems and opportunities of a particular moment are and how to move forward in the next possibly quite small steps. Again, this requires awareness: awareness of the particular health situation, disease susceptibilities, and vulnerabilities of the individual at that particular moment, what is happening in science and opportunities for new interventions that may be opening up, possibilities for modifying daily routines or simple processes such as related to movement or what to eat when. What can be very helpful is mobilizing a strong supporting external network of people to assist in this process.
Where do other scientists stand in the kinds of views expressed here?
I think there is widespread agreement to many of the central points made here. See for example the recent science news story Strategy proposed for preventing diseases of aging/ “Researchers argue that medicine focuses too much on fighting diseases individually instead of concentrating on interventions that prevent multiple chronic diseases and extend healthy lifespan. They call for moving forward with strategies that have been shown to delay aging in animals. In addition to promoting a healthy diet and regular exercise, these strategies include manipulating molecular pathways that slow aging and promote healthy longevity. — “Heart failure doesn’t happen all at once,” Fontana said. “It takes 30 or 40 years of an unhealthy lifestyle and activation of aging-related pathways from metabolic abnormalities such as high blood pressure, high cholesterol and type 2 diabetes to give a person heart failure in his 60s. So we propose using lifestyle interventions — such as a personalized healthy diet and exercise program — to down-regulate aging pathways so the patient avoids heart failure in the first place. — Fontana and his colleagues write that most clinicians don’t realize how much already is understood about the molecular mechanisms of aging and their link to chronic diseases. — But challenges abound. The most important change, they argue, is in mindset. ”
Wrapping it up
I have outlined a general process philosophy for creating personal health and longevity which happens to be the one I have been seeking to pursue for a number of years now. I outlined the similarity of this philosophy to that of TMQ, an industrial philosophy which over a 60 year interval has allowed us to effectively quadruple the lifespan of automobiles. I speculate on how through applying TMQ-like principles we might go as far as doubling our expected human lifespans in as little as 30 years. If I were asked “How strongly do you feel about this incremental approach, Vince?” my answer is “I have been betting my life on it.”
By Jim Watson with editorial and minor contributions by Vince Giuliano
Much has been learned over the years about Alzheimer’s Disease (AD) and there are several sides to the unfolding story which considered together are making more and more sense. This blog entry focuses on the role of amyloid beta in the brain, a major factor universally acknowledged to be of importance – although in the absence of agreement as to exactly what that importance is. We advance the hypothesis that a major factor in the progression of the disease is failure of clearance of soluble amyloid beta in the brain. Further, we believe that this is due to lack of adequate binding of APOE species to the LDL receptors. The issue can be some combination of APOE gene polymorphisms which are well known to create AD susceptibly and age-related insufficient production in the brain of the LDL receptor related protein (LRP).
We also discuss several related aspects of the AD story including why presence of beta amyloid is only a possible but not a definitive indicator of AD, details related to how beta amloid can or cannot be created via secretases, the beta-amyloid clearance role of the LDL receptor, age-related inflammaging, the role of the RAGE receptor and AB neurotoxicity, why AB-42 is much more dangerous than AB-40, roles of microglia, how beta-amloid kills neurons and when it does not do that, the roles of APOE and its polymorphisms, and two new high accuracy tests for predicting whether AD is likely to develop..
First, we recapitulate some background on what beta-amyloid is and is is typically involved in cases of AD. The hypothysis that extra-cellular beta amyloid deposits in the brain is usually a key early indicator of AD goes back to the early 90s and has now become quite mainline. The 2002 document The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeuticsrelated “It has been more than 10 years since it was first proposed that the neurodegeneration in Alzheimer’s disease (AD) may be caused by deposition of amyloid beta-peptide (Abeta) in plaques in brain tissue. According to the amyloid hypothesis, accumulation of Abeta in the brain is the primary influence driving AD pathogenesis. The rest of the disease process, including formation of neurofibrillary tangles containing tau protein, is proposed to result from an imbalance between Abeta production and Abeta clearance.” This is still a good formulation of the hypothysis. Problems are, of course, 1. Some people develop high levels of beta-amloid but never progress to showing dementia or AD, and 2. Some people develop dementia thought to be AD but do not have high levels of brain beta-amyloid. In any case for our purposes, if amloid-beta is involved in AD, it is responsible for early stages of the pathology – ones leading to the tau tangles characteristic of later stages.
As time progressed a more nuanced view of the amloid hypothysis has emerged such as laid out in the 2009 publication Reassessing the Amyloid Cascade Hypothesis of Alzheimer’s Disease: “Since its inception, the amyloid cascade hypothesis has dominated the field of Alzheimer’s disease (AD) research and has provided the intellectual framework for therapeutic intervention. Although the details of the hypothesis continue to evolve, its core principle has remained essentially unaltered. It posits that the amyloid-β peptides, derived from amyloid precursor protein (APP), are the root cause of AD. Substantial genetic and biochemical data support this view, and yet a number of findings also run contrary to its tenets. The presence of familial AD mutations in APP and presenilins, demonstration of Aβ toxicity, and studies in mouse models of AD all support the hypothesis, whereas the presence of Aβ plaques in normal individuals, the uncertain nature of the pathogenic Aβ species, and repeated disappointments with Aβ-centered therapeutic trials are inconsistent with the hypothesis. The current state of knowledge does not prove nor disprove the amyloid hypothesis, but rather points to the need for its reassessment. A view that Aβ is one of the factors, as opposed to the factor, that causes AD is more consistent with the present knowledge, and is more likely to promote comprehensive and effective therapeutic strategies.”
SOME KEY PIECES OF THE BETA AMYLOID JIGSAW PUZZLE
As I see it, all the pieces of the Alzheimer’s puzzle are finally starting to fit together. I will try to describe key piecea of the puzzle using some easy-to-understand analogies. Each item below is a piece of the interlocking puzzle. Each piece has at least 4 sides to it as follows:
1. Dispelling the Myth about “Amyloid negative Alzheimer’s disease”
Although the correlation between Amyloid beta accumulation and Dementia is far from perfect, there is no question that Amyloid beta accumulates in over 95% of patients who develop age-related dementia. Today we know that many other distinctly different causes of dementia exist that can present with the exact same signs and symptoms of AD.
As a result, today it is believed that cases of dementia where no Amyloid beta is present on autopsy, one of the following causes of the dementia explain the “Amyloid beta Negative dementia”:
Vascular dementia
Lewy body disease
Chronic traumatic encephalopathy (CTE)
Frontotemporal dementia
Parkinson’s disease with dementia (15% of PD)
Amyotrophic lateral sclerosis (ALS) with dementia
The Misinterpreted Nun Study
Digging a little deeper into the inconclusive relationship of beta amyloid to AD, we go on to dispell the myth of “Amyloid negative Alzheimer’s disease” and provide an explanation for for the Nuns who had Amyloid beta on autopsy, but had no Alzheimer’s disease. (Aka. A slice of the Venn Diagram of Dementia). Our eventual purpose of this blog entry is to lay out what we strongly suspect may be happening.
There is a famous “Nun Study” reporting on autopsies done on a series of Catholic Nuns who died. Some had Alzheimer’s disease but many died with no dementia. Although most of the autopsies of Nuns with dementia showed Amyloid beta, some did not. What was even more surprising was that some Nuns with no dementia had amyloid beta accumulation. This study has been often quoted by those who do not believe that amyloid beta causes Alzheimer’s disease. In many ways, this is analogous to the debate of cholesterol and arteriosclerosis. Today most experts do not believe that arteriosclerosis is a “cholesterol disease”. Instead, it is a “cholesterol transport disease” (aka a lipoprotein clearance disease). Likewise, most experts believe that Alzheimer’s is NOT an Amyloid beta disease but a “Amyloid beta clearance disease.” As discussed below, AD may be consequential to possible problems with lipoprotein E and/or the LDL receptor related protein that it binds to for clearing amyloid beta.
Just like the presence of cholesterol does not mean you have arteriosclerosis, the presence of amyloid beta does not mean you have Alzheimer’s disease. This is a key point that most everyone misunderstands about these two diseases. What is even more surprising is how the two “puzzles” (arteriosclerosis and Alzheimer’s) have a common puzzle piece – the LDL receptor! “The Low-Density Lipoprotein (LDL) Receptor is a mosaic protein of 839 amino acids (after removal of 21-amino acid signal peptides)[1] that mediates the endocytosisof cholesterol-rich LDL. It is a cell-surface receptor that recognizes the apoprotein B100, which is embedded in the outer phospholipid layer of LDL particles. The receptor also recognizes the apoE protein found in chylomicron remnants and VLDL remnants (IDL). In humans, the LDL receptor protein is encoded by the LDLR gene.[2][3] It belongs to the Low density lipoprotein receptor gene family[4](ref). ” The LDL receptor on the surface of macrophages in the blood vessel wall and the LDL receptor related protein on the Brain’s microglial cells are almost exactly the same and when these receptors are absent (or decline with aging), there is a 100% incidence of arteriosclerosis and Alzheimer’s disease. Thus many have suggested that we call arteriosclerosis and Alzheimer’s disease both by the same name – “LDL Receptor Disease“. However this is a little bit of an oversimplification, as you will see below.
A person with clinical dementia may not have Alzheimer’s disease, but may have one of the other causes of dementia. This explains “amyloid beta negative Dementia” seen in the famous “Nun study”.
The other part of the “Nun Study”Is also explainable – i.e. the Nuns with no dementia who had Amyloid beta accumulation. This I believe is due to the body’s built in cellular stress coping mechanisms controlled by “gene switches” — transcription factors. The Transcription factors that prevented these Nuns from developing Alzheimer’s disease are named FoxO3a, FoxO1, Nrf2, COREST, PGC-1a, and many other hard-to-understand names. These “amyloid beta positive Nuns” had no dementia because they had developed strong cellular stress resistance. Their neurons were resistant to cell death. This effect is an aspect of hormesis, a non-linear response to stress often discussed in this blog. “. Others like to call it “ischemic preconditioning”. Others explain this phenomena by the molecular mechanisms involved with caloric restriction. (T hey are all different views of the same cellular stress resistance system). Regardless of the terminology used to describe cellular stress resistance, there is no debate that you can reduce cell death by making your brain cells resistant to cell stress.
So, amyloid beta is a “cellular stressor” that can either kill your neurons (and you will develop dementia) or you can become resistant to Amyloid beta toxicity on a cellular level. With aging, everyone develops multiple types of amyloidosis. After age 110, the transthyretin amyloid protein builds up. Are you going to die of “amyloid stress” or are you going to become resistant to”amyloid stress”. This is the question that Hamlet asked himself In the famous Shakesphere line..”to be or not to be.” I would Rephrase this: “to die of amyloid stress or become resistant to Amyloid stress….this is the question. ”
If you grow old enough, there will be Amyloid beta fibrils present on your autopsy after death. However, your neurons do NOT need to be dead when you die. This explains the “dementia-free, Abeta positive Nuns”.
To avoid mislabeling the cause of dementia, the confirmatory diagnosis of Alzheimer’s disease requires an autopsy that shows histologic evidence of Amyloid beta presenting 8 nm aggregates. This is typically referred to as “amyloid Beta fibrils”. If this is not present, then another cause for the dementia is entertained.
2. Key role of the LDL receptor protein in beta amyloid clearance
We asserted above that AD is involved with inadequate clearance of soluble amyloid beta in the brain. LRP binds to three ligands (ApoE, alpha2 macroglobin, and KPI), two of which have been shown to bind to both AB-40 and AB-42. Both ApoE and alpha2 macroglobulin (a2M) bind to Abeta. Then the ApoE/ABeta or a2M/Abeta binds to the LRP and is endocytosed. This has been known for 14 years, which is somewhat shocking! See the 2000 article Modulation of amyloid β-protein clearance and Alzheimer’s disease susceptibility by the LDL receptor–related protein pathway. “Susceptibility to Alzheimer’s disease (AD) is governed by multiple genetic factors. Remarkably, the LDL receptor–related protein (LRP) and its ligands, apoE and α2M, are all genetically associated with AD. In this study, we provide evidence for the involvement of the LRP pathway in amyloid deposition through sequestration and removal of soluble amyloid β-protein (Aβ). We demonstrate in vitro that LRP mediates the clearance of both Aβ40 and Aβ42 through a bona fide receptor-mediated uptake mechanism. In vivo, reduced LRP expression is associated with LRP genotypes and is correlated with enhanced soluble Aβ levels and amyloid deposition. Although LRP has been proposed to be a clearance pathway for Aβ, this work provides the first in vivo evidence that the LRP pathway may modulate Aβ deposition and AD susceptibility by regulating the removal of soluble Aβ.” –
“Discussion: The genetic associations of APOE, α2M, and LRP to late-onset AD are particularly intriguing in light of the fact that both apoE and α2M are two key ligands for LRP (5, 6, 8). Both apoE and α2M avidly bind Aβ in vitro and in vivo (5, 35). These observations, together with the finding that LRP and all of its ligands are present in senile plaques (7), strongly implicate the pathogenic importance of the LRP pathway in AD. We found that LRP levels are significantly reduced in AD, compared with healthy controls. Linear-regression analysis revealed that LRP levels progressively decline with the increasing age of control subjects (an inverse correlation) and are further reduced in AD subjects. Among AD patients, however, increased LRP levels were correlated with later age of disease onset, indicating that higher LRP levels might be protective against AD. This apparent protective effect was accentuated among noncarriers of the APOE ε4 allele. As increasing age is the primary risk factor for AD, these data indicate that reduced LRP expression may be one factor contributing to AD susceptibility. This notion is highly consistent with the negative association of the LRP T allele to AD (8, 9, 11, 12) and increased LRP levels among T-allele carriers demonstrated in this study. Although the biological mode of the LRP T allele requires further characterization, we hypothesize that the C766T polymorphism is in linkage disequilibrium with a causative mutation or polymorphism that regulates LRP expression (for example, promoter/enhancer) in the aging brain. ‘’
“In the current study, we provide compelling evidence that Aβ uptake via the α2M-LRP pathway is through a bona fide receptor-mediated mechanism and not through nonspecific Aβ degradation or fluid-phase pinocytosis. –“
“The results of the current study provided the first in vivo evidence of the LRP-clearance pathway in AD pathogenesis. Our observations lead us to postulate that reduced LRP expression is a contributing risk factor for AD, possibly by impeding clearance of soluble Aβ complexes. Functional characterization of α2M polymorphisms associated with AD (6) and future transgenic animal models of LRP and/or α2M expression should further elucidate the mechanism of Aβ clearance and AD pathogenesis. The observation that AD risk or protection associated with LRP levels is strongest among noncarriers of the APOE ε4 allele is particularly interesting in light of the ligand/receptor relationship between apoE and LRP. Because the receptor function of LRP obviously depends on intact activity of its ligands, we hypothesize that high levels of LRP cannot effectively rescue the pathogenic effects of apoE4, the latter operating at a step that negates the clearance mechanism. However, in the presence of apoE2 or apoE3, where the ligand complexes are not perturbed, alterations in LRP level and, in turn, clearance activity become highly consequential for AD pathogenesis. Since increased LRP expression may promote both neuronal survival mediated by apoE2 and apoE3 isoforms and also enhance the clearance of soluble Aβ complexes, the current data provide an alternative direction for AD therapeutic intervention by targeting the Aβ/LRP clearance pathway in non-APOE ε4 carriers.”
Amyloid beta is made up of the cleaved “tail” of the Amyloid precursor protein (APP). APP is a plasma membrane protein that Requires a “haircut”, once APP Trans locates from the ER (protein factory) to the plasma membrane.
The “barbers” that give APP a “haircut” are called “Secretases”. Three “barbers” exist that cut hair at different lengths. The pieces of “hair” cut off APP are the “tails” of the APP molecule and are what Amyloid beta is made up of.
3. It takes two Barbers to cut hair In the brain – each hair must be cut twice!
It takes two barbers to cut the hair on the APP protein. If alpha secretase “cuts first” and gamma secretase cuts APP second, no amyloid accumulates in the brain. If BACE “cuts first” and gamma secretase cuts second, then the piece that is produced forms Amyloid aggregates with lengths of 17 to 42 amino acids. The longer pieces of “cut hair” left behind outside of the cell are called Amyloid beta 40 (AB-40) and amyloid beta 42 (Ab-42).
AlthoughAB-42 is found in much smaller amounts, compared to the more common AB-40, the AB-42 piece of “cut hair” is much more toxic. The next piece of the puzzle explains why.
4. Why Amyloid beta aggregates: “The salt bridge between Asp23 and Lys28″
Like all polypeptides, Amyloid beta has amino acids with positive and negatively charged side chains. One negatively charged side chain (on aspartate, #23) and one positively charged side chain (on lysine, #28) are attracted to each other and form a “salt bridge”. This salt bridge between Asp23 and Lys28 forces the chain to form a “Hairpin turn,” looping back on itself. (A salt bridge is a bond between a positively charged amino acid side chain and a negatively charged amino acid side chain). The hairpin shape of Abeta makes multiple copies of the polypeptide stick together in parallel, forming a 3D structure called a “beta Pleated sheet”. See the following diagram. This beta-pleated sheet structure is what is referred to as a “Amyloid fibril” and is about 8 nm in size. It is not soluble and starts aggregating outside of the cell.
Amyloid beta 40 is the common, more soluble form of A-beta. A-beta 42 has two more amino acids on the tail end of the peptide. One of these, amino acid #42, is an alanine, which can loop back to form a salt bridge with amino acid #35, methionine (Met35). This extra hairpin turn of AB-42 makes it less soluble and more toxic. The toxicity of AB-42 is much greater than AB-40. One possible reason for this is described below.
“(a) Sequence of Aβ42 that is derived from human APP. (b) Structural constraints in Aβ40 and Aβ42 fibrils. NMR measurements of Aβ40 fibrils have shown that residues 1–10 are unstructured and residues 11–40 adopt a β-turn-β fold19, 20. Side chain packing is observed between Phe19 and Ile32, Leu34 andVal36 and between Gln15 and Val36 as well as between His13 and Val40 (blue dashed lines). In Aβ42 fibrils, residues 1–17 may be unstructured (in gray), with residues 18–42 forming a β-turn-β fold14. Molecular contacts have been reported within the monomer unit of Aβ42 fibrils between Phe19 and Gly38 (red dashed line)14 and between Met35 and Ala42 (orange dashed line)18. In both Aβ40 and Aβ42, the turn conformation is stabilized by hydrophobic interactions (green residues) and by a salt bridge between Asp23 and Lys28 (black dashed line).”
6. Direct neurotoxicity of Abeta
“The mechanism by which amyloid beta may damage and kill neurons is by generating reactive oxygen species during the process of its self-aggregation. When this occurs on the membrane of neurons it causes lipid peroxidation and the generation of a toxic aldehyde called 4-hydroxynonenal which, in turn, impairs the function of ion-motive ATPases, glucose transporters and glutamate transporters. As a result, amyloid beta promotes depolarization of the synaptic membrane, excessive calcium influx and mitochondrial impairment.[51](ref Wikipedia).
“Amyloid-beta(Aβ or Abeta) denotes peptidesof 36–43 amino acidsthat are crucially involved in Alzheimer’s diseaseas the main component of the amyloid plaquesfound in the brains of Alzheimer patients. The peptides result from the amyloid precursor protein(APP), which is being cut by certain enzymes to yield Aβ. Aβ molecules can aggregate to form flexible soluble oligomerswhich may exist in several forms. It is now believed that certain misfolded oligomers (known as “seeds”) can induce other Aβ molecules to also take the misfolded oligomeric form, leading to a chain reaction akin to a prioninfection. The seeds or the resulting amyloid plaques are toxic to nerve cells. The other protein implicated in Alzheimer’s disease, tau protein, also forms such prion-like misfolded oligomers, and there is some evidence that misfolded Aβ can induce tau to misfold.[2] — A recent study suggested that APP and its amyloid potential is of ancient origins, dating as far back as early deuterostomes.[3] (ref Wikipedia).”
7. Abeta and RAGE receptor mediated neurotoxicity: The story of NF-kB, MAPK, and “Inflammaging”
There is a pro-inflammatory membrane receptor found on all cells that binds to “Advanced Glycation End products”, aka AGEs. Hemoglobin A1c is an example of an AGE. The Membrane receptor for AGEs is called the “Receptor for Advanced Glycation Endproducts” or RAGE. Amyloid beta binds to the RAGE receptor and triggers sustained activation if pro-inflammatory pathways including the NF-kB and MAPK pathways. This is especially bad because it induces a prolonged activation of these pathways, whereas most stimuli induce a temporary, short period of pro-inflammatory gene expression.
Although other pathways account for the destruction of Abeta (such as the LDL receptor related protein, or LRP), the RAGE-mediated activation of inflammation in neurons, astrocytes, and glial cells is thought to induce cellular senescence, “inflammaging”, and neuronal cell death by multiple gene products induced by NF- kB and MAPK, such as IL-1b, TNF-a, IL-6, IL-8, ROS, RNS, and other Molecules secreted as part of the SASP. In all of this “inflammaging,” autophagy is also up regulated. Eventually the neurons die by apoptosis or autophagy-induced Cell death (AICD). .
Ref: RAGE and amyloid-beta peptide neurotoxicity in Alzheimer’s disease. “Amyloid-beta peptide is central to the pathology of Alzheimer’s disease, because it is neurotoxic–directly by inducing oxidant stress, and indirectly by activating microglia. A specific cell-surface acceptor site that could focus its effects on target cells has been postulated but not identified. Here we present evidence that the ‘receptor for advanced glycation end products’ (RAGE) is such a receptor, and that it mediates effects of the peptide on neurons and microglia. Increased expressing of RAGE in Alzheimer’s disease brain indicates that it is relevant to the pathogenesis of neuronal dysfunction and death). This topic is discussed in the earlier-mentioned blog entries on microglial activation and neurological diseases.
Site-Specific Blockade of RAGE-Vd Prevents Amyloid-β Oligomer Neurotoxicity. “In the genesis of Alzheimer’s disease (AD), converging lines of evidence suggest that amyloid-β peptide (Aβ) triggers a pathogenic cascade leading to neuronal loss. It was long assumed that Aβ had to be assembled into extracellular amyloid fibrils or aggregates to exert its cytotoxic effects. Over the past decade, characterization of soluble oligomeric Aβ species in the brains of AD patients and in transgenic models has raised the possibility that different conformations of Aβ may contribute to AD pathology via different mechanisms. The receptor for advanced glycation end products (RAGE), a member of the Ig superfamily, is a cellular binding site for Aβ. Here, we investigate the role of RAGE in apoptosis induced by distinct well characterized Aβ conformations: Aβ oligomers (AβOs), Aβ fibrils (AβFs), and Aβ aggregates (AβAs). In our in vitrosystem, treatment with polyclonal anti-RAGE antibodies significantly improves SHSY-5Y cell and neuronal survival exposed to either AβOs or AβAs but does not affect AβF toxicity. Interestingly, using site-specific antibodies, we demonstrate that targeting of the Vd domain of RAGE attenuates AβO-induced toxicity in both SHSY-5Y cells and rat cortical neurons, whereas inhibition of AβA-induced apoptosis requires the neutralization of the C1d domain of the receptor. Thus, our data indicate that distinct regions of RAGE are involved in Aβ-induced cellular and neuronal toxicity with respect to the Aβ aggregation state, and they suggest the blockage of particular sites of the receptor as a potential therapeutic strategy to attenuate neuronal death.”)
Involvement of microglial receptor for advanced glycation endproducts (RAGE) in Alzheimer’s disease: identification of a cellular activation mechanism. “Receptor-mediated interactions with amyloid beta-peptide (Abeta) could be important in the evolution of the inflammatory processes and cellular dysfunction that are prominent in Alzheimer’s disease (AD) pathology. One candidate receptor is the receptor for advanced glycation endproducts (RAGE), which can bind Abeta and transduce signals leading to cellular activation. Data are presented showing a potential mechanism for Abeta activation of microglia that could be mediated by RAGE and macrophage colony-stimulating factor (M-CSF). Using brain tissue from AD and nondemented (ND) individuals, RAGE expression was shown to be present on microglia and neurons of the hippocampus, entorhinal cortex, and superior frontal gyrus. The presence of increased numbers of RAGE-immunoreactive microglia in AD led us to further analyze RAGE-related properties of these cells cultured from AD and ND brains. Direct addition of Abeta(1-42) to the microglia increased their expression of M-CSF. This effect was significantly greater in microglia derived from AD brains compared to those from ND brains. Increased M-CSF secretion was also demonstrated using a cell culture model of plaques whereby microglia were cultured in wells containing focal deposits of immobilized Abeta(1-42). In each case, the Abeta stimulation of M-CSF secretion was significantly blocked by treatment of cultures with anti-RAGE F(ab’)2. Treatment of microglia with anti-RAGE F(ab’)2 also inhibited the chemotactic response of microglia toward Abeta(1-42). Finally, incubation of microglia with M-CSF and Abeta increased expression of RAGE mRNA. These microglia also expressed M-CSF receptor mRNA. These data suggest a positive feedback loop in which Abeta-RAGE-mediated microglial activation enhances expression of M-CSF and RAGE, possibly initiating an ascending spiral of cellular activation.”)
Advanced glycation endproducts and their receptor RAGE in Alzheimer’s disease “In this review, we revisit the hypothesis that advanced glycation endproducts (AGEs) and their receptor RAGE may play an important role in disease pathogenesis. Accumulation of AGEs in cells and tissues is a normal feature of aging, but is accelerated in AD. In AD, AGEs can be detected in pathological deposits such as amyloid plaques and neurofibrillary tangles. AGEs explain many of the neuropathological and biochemical features of AD such as extensive protein crosslinking, glial induction of oxidative stress and neuronal cell death. Oxidative stress and AGEs initiate a positive feedback loop, where normal age-related changes develop into a pathophysiological cascade. RAGE and its decoy receptor soluble RAGE, may contribute to or protect against AD pathogenesis by influencing transport of β-amyloid into the brain or by manipulating inflammatory mechanisms. Targeted pharmacological interventions using AGE-inhibitors, RAGE-antagonists, RAGE-antibodies, soluble RAGE or RAGE signalling inhibitors such as membrane-permeable antioxidants may be promising therapeutic strategies to slow down the progression of AD.”
Amyloid beta clearance – the role of ApoE as the “1st clean up protein”
ApoE is a “transport protein” whose primary function in the brain is to help transport Cholesterol and phospholipids in the brain to the sites of new neuron spine formation (called neurites), new neuron cell growth, and myelin formation. The role of ApoE in this transport system is that of a “docking protein”, much like ApoA and ApoB are docking proteins for the HDL and LDL transport systems in the blood that transport cholesterol to the liver (HDL) and away from the liver (LDL).
ApoE has an additional “2nd job” in the brain, however. It is also part if the “clean up crew” for clearing soluble Amyloid beta out if the brain. ApoE first sticks to the hydrophobic portions of the Amyloid beta and then clears the Abeta out of the extra cellular space by binding to the LDL receptor related protein (LRP) on the surface of the brain microglial cells. The reason ApoE4 is associated with an increased risk of AD is explained below in section 10.
9. Abeta clearance by alpha2 macro-globin (a2M) – the 2nd “clean up Protein
There is a second clean-up system for disposing of extracellular junk proteins. This system utilizes a protein called “alpha 2 macro-globin, or a2M. a2M binds to soluble amyloid beta and then binds to LRP, just like ApoE.
10. ApoE4 vs ApoE3 vs ApoE2
Three variants of the ApoE protein are found in the population at large. They vary in sequence at two locations on the ApoE amino acid chain. The 3 forms of the protein are called ApoE2, ApoE3, and ApoE4. ApoE4 has a different binding Affinity for LRP than ApoE3 or ApoE2. This is why ApoE4 does not clear the Amyloid beta as well.
The 2000 publication Modulation of amyloid β-protein clearance and Alzheimer’s disease susceptibility by the LDL receptor–related protein pathway reports: “Susceptibility to Alzheimer’s disease (AD) is governed by multiple genetic factors. Remarkably, the LDL receptor–related protein (LRP) and its ligands, apoE and α2M, are all genetically associated with AD. In this study, we provide evidence for the involvement of the LRP pathway in amyloid deposition through sequestration and removal of soluble amyloid β-protein (Aβ). We demonstrate in vitro that LRP mediates the clearance of both Aβ40 and Aβ42 through a bona fide receptor-mediated uptake mechanism. In vivo, reduced LRP expression is associated with LRP genotypes and is correlated with enhanced soluble Aβ levels and amyloid deposition. Although LRP has been proposed to be a clearance pathway for Aβ, this work provides the first in vivo evidence that the LRP pathway may modulate Aβ deposition and AD susceptibility by regulating the removal of soluble Aβ.”
– “The LDL receptor–related protein (LRP) is a multifunctional receptor that mediates the internalization and degradation of ligands involved in metabolic pathways of lipoproteins and protease/protease-inhibitor complexes (1), including α2-macroglobulin (α2M) (2), apoE (3), and Kunitz protease inhibitor (KPI) containing forms of amyloid precursor protein (APP) (4). Remarkably, the aforementioned ligands are all genetically associated with Alzheimer’s disease (AD) (5, 6) and are found in senile plaques in brains of AD patients (7). To date, the strongest evidence directly implicating a role of LRP in AD is from genetic studies first reported by us (8) and subsequently confirmed in four independent case-control cohorts (9–12). In addition, another genetic polymorphism in LRP was found to be associated with AD (13), further evidence corroborating the LRP gene as an important AD-susceptibility locus. Our study reported a genetic polymorphism (C766T) in exon 3 of LRP that is under-represented in AD and associated with later age of disease onset. However, the underlying biologic relevance of the silent LRP C766T polymorphism is unclear. Moreover, the precise mechanisms by which LRP and its ligands may contribute to AD pathogenesis are unknown.”
– “In the current study, we provide compelling evidence that Aβ uptake via the α2M-LRP pathway is through a bona fide receptor-mediated mechanism and not through nonspecific Aβ degradation or fluid-phase pinocytosis. This was shown by the competition of 125I-Aβ uptake with excess unlabeled Aβ complexes and the complete saturation of 125I-Aβ uptake at physiological concentrations. So far, no other Aβ uptake pathway meets the criteria for a bona fide receptor-mediated mechanism. Although the scavenger receptor has been postulated to mediate the uptake of amyloid fibrils, such process is not subject to competition and saturation of the receptor (30, 31). At another level of Aβ catabolism, recent observations have indicated that neutral endopeptidase and insulin-degrading enzyme are both capable of degrading extracellular Aβ in a cell-autonomous manner (36,37). Thus, it is likely that there are multiple ways of mediating Aβ degradation in vivo. Our study demonstrated that LRP does not alter the secretion of Aβ from APP695-expressing cells but is required for α2M-mediated clearance of soluble Aβ. Because both LRP and APP695 are predominantly expressed in neurons, reduced LRP levels in the AD brain is predicted to negatively impact the clearance of soluble Aβ but not its production in neurons. However, it is important to note that LRP overexpression in LRP-deficient CHO cells results in altered trafficking of KPI containing APP751 (28), possibly through the APP-LRP physical interaction (4). Thus, it is possible that LRP also alters APP trafficking (i.e., internalization/recycling) and Aβ generation through other mechanisms. As APP isoforms are differentially expressed in neurons and glia, LRP-clearance activity versus altered APP trafficking might be differentially modulated across cell types.”
– “The genetic association of LRP T polymorphism with both increased LRP expression and reduced amyloid deposition is intriguing in light of the in vitro evidence of Aβ clearance through the LRP pathway (16, 17). These observations are now further strengthened by genetic association of LRP with soluble Aβ levels in plasma. It is noteworthy that the pathogenic Aβ42 species is as effectively cleared through the LRP pathway as Aβ40 (Figure 1a), an activity that may dramatically impact amyloid deposition in vivo. Accordingly, we interpret these data to indicate that reduced LRP expression, at least in part, contributes to increased Aβ levels and amyloid deposition by negatively impacting Aβ clearance. This interpretation is consistent with our observation that reduced LRP expression is also correlated with increased AD susceptibility and earlier age of disease onset. In our cell-culture system, we demonstrated the requirement of LRP in the α2M-mediated clearance of Aβ. However, it has been reported that apoE and lactoferrin, two other LRP ligands, also sequester Aβ and mediate its clearance (16, 38). Thus, reduced LRP levels may impede the clearance of various Aβ complexes. Interestingly, many LRP ligands, including apoE, α2M, and lactoferrin, are produced from astrocytes, whereas LRP is largely expressed in neurons. Thus, it is likely that receptor-mediated uptake and clearance of soluble Aβ complexes occur in neurons, whereas uptake of fibrillar amyloid is mediated by microglia (30, 31). In this regard, downregulation of LRP expression has been linked to proinflammatory stimuli such as LPS and IFN-γ in cultured cells (39, 40). We speculate that proinflammatory processes present in the AD brain may induce downregulation of LRP expression, further reducing Aβ clearance and enhancing amyloid deposition. Since LRP mediates the normal function of neuronal remodeling through internalization of apoE (19), reduced LRP expression in aging and disease may also compromise neuronal viability independent of the effects on Aβ clearance.”
– “The results of the current study provided the first in vivo evidence of the LRP-clearance pathway in AD pathogenesis. Our observations lead us to postulate that reduced LRP expression is a contributing risk factor for AD, possibly by impeding clearance of soluble Aβ complexes.”
New predictive tests for Alzheimer’s Disease
Over the years a number of tests have been proposed or developed for confirming the existence of AD or predicting the onset of AD in presymptomatic individuals, These tests have ended to be expensive imaging procedures (e.g. PET scan, 3D volumetric MRI, etc), or require invasive procedures (e.g. a CSF test which requires a lumbar puncture) or have relatively low accuracy (e.g. like some of the individual tests for proteins developed in the 1990s and early 2000s mentioned below).
Lipid biomarker test
A lipid biomarker test for AD has been under development at Georgetown and Rochester Universities. As reported in the 2014 story in the ALZFORUM Do Lipids Hold the Key to Blood-Based Alzheimer’s Test? “Researchers have long been searching for a blood-based test that could predict Alzheimer’s disease. In the March 9 Nature Medicine, researchers led by Howard Federoff at Georgetown University Medical Center, Washington, D.C., and Mark Mapstone at the University of Rochester School of Medicine, New York, report that a panel of 10 lipids could one day fit the bill. In a small study, plasma levels of these lipids distinguished, with 90 percent accuracy, who would develop cognitive impairment over the next two to three years. If the results are confirmed in larger samples, it would encourage hopes for a blood test for Alzheimer’s. However, the authors stress that is still years away. Numerous questions remain, including whether the observed lipid changes are specific to AD, or represent a more general marker for neurodegeneration.”
“The ability to identify individuals who are at risk of developing Alzheimer’s before the clinical manifestation of cognitive impairment has long been a Holy Grail of the neuromedicine community,” said Mark Mapstone, Ph.D., a neuropsychologist with the University of Rochester School of Medicine and Dentistry and lead author of the study.“Current efforts to develop a treatment for this disease are coming up short because they are probably being used too late. Biomarkers that can allow us to intervene early in the course of the disease could be a game-changer(ref).” Other criteriia for an effective predictive test for AD is that is should be relatively cheap and non-invasive It is expected that this “lipid bio marker test” will cost around $200.
Combination protein test
Lipids are tricky to measure in blood, however. Proteins, on the other hand, are much easier to measure. A new “protein test” from the European Consortium group looks at 10 plasma proteins, rather than 10 lipids. It has an accuracy of 87% and a specificity of 88%. (Not quite as good, but the test may be easier to do in the lab. The test is described in the July 2014 publication Plasma proteins predict conversion to dementia from prodromal disease. “Background:The study aimed to validate previously discovered plasma biomarkers associated with AD, using a design based on imaging measures as surrogate for disease severity and assess their prognostic value in predicting conversion to dementia. – Methods: Three multicenter cohorts of cognitively healthy elderly, mild cognitive impairment (MCI), and AD participants with standardized clinical assessments and structural neuroimaging measures were used. Twenty-six candidate proteins were quantified in 1148 subjects using multiplex (xMAP) assays. Results: Sixteen proteins correlated with disease severity and cognitive decline. Strongest associations were in the MCI group with a panel of 10 proteins predicting progression to AD (accuracy 87%, sensitivity 85%, and specificity 88%). Conclusions: We have identified 10 plasma proteins strongly associated with disease severity and disease progression. Such markers may be useful for patient selection for clinical trials and assessment of patients with predisease subjective memory complaints.”
Here is the list of 10 proteins from this new “protein test” and my comments on each one, along with some references. It is surprising how many are actually “neuroprotective Proteins” (5), rather than “neurotoxic proteins” (3) and that many have been known for some time.
1. TTR – Transthyretin is a protein that binds to amyloid beta and prevents it from aggregating. Thus it is neuroprotective. In AD, there is lower levels of TTR.
2. Clusterin (aka ApoJ) – This is another “clean up crew” protein that cleans up bad (Damaged) lipids in the cell membranes. It is neuroprotective in Alzheimer’s disease.
3. Cystatin C – This is the most abundant serine protease inhibitor found in the extra-cellular space. It binds to amyloid beta and prevents it from aggregating and forming sheets of amyloid. Thus, it is Neuroprotective.
4. Alpha 1 acid Glycoprotein – This is an acute phase protein, like C-reactive protein. It is produced in response to inflammation. In sick people in the hospital, it predicts death.
5. ICAM1 – This is an intercellular adhesion molecule. It attracts inflammatory cells such as macrophages to the site and holds the cells there (I.e. Makes the site “sticky”). Levels in the brain are increased with AD and it appears to be coming from astrocytes,
9. RANTES (ApoC3) – This appears to be a “cellular stress resistance protein”. When it is secreted in cases of AD, it protects the nerve cells from dying
10. ApoE genotype – This is obviously a good predictor of earlier onset of LOAD.
More on Alzheimer’s disease
There is much more to the AD story besides the beta-amloid aspects and others discussed here. Jim has more to say. previous blog entries are mentioned above, and Vince has been working for some time on a blog on the tau tangles aspects of AD and other neurological diseases. So, stand by for more!
Immune system functionality declines significantly with advancing age, leading to increased susceptibility to infectious and many other diseases. This is a very important aspect of aging as we know it. This blog entry looks again at the science related this decline and to recent research that may lead tof intervention for halting or reversing it.
BACKGROUND
In large part, this blog entry picks up from the September 2011 blog entry Aging and the immune system – focus on naïve T-cells. If you are not familiar with the topic, we suggest you start by reading that blog before proceeding further here. A central focus of that blog was reversal of thymic involution – age related shrinking and decline of functionality of the thymus gland, This gland makes T cells that are necessary for the operation of the adaptive immune system.
We have two interacting immune systems, the innate immune system which is older and common to more primitive species, a system that offers fixed responses to pathogens and our adaptive immune system which consists of a system of specialized cells and processes that remembers pathogens we encountered earlier and life to enable a rapid and effective defensive response if they are encountered again even years afterward. The 2011 blog focuses on the adaptive immune system and introduces the main cellular actors related to it including T cells, B cells, naïve cells, memory cells, effector cells and thymocytes. It also describes the key role of the thymus gland early in life for processing thymocytes into mature naïve T cells – a role that drops off rapidly after adolescence and is virtually non-existent in older people.” And it covers some possible approaches for regenerating the thymus gland. This present blog covers additional background and more-recent research and insights applicable to that same issue.
MAJOR BOTTOM LINES OF THIS BLOG ENTRY
This blog contains a great deal of detailed information, and it is easy to lose sight of the most important conclusions. So, we list them here.
Many factors are involved factors in age-related decline in the functioning of the immune systems. Among these is the involution and gradual disappearance of the thymus gland which is key for generating new T cells. But this is only one of several age-related processes related to decline in immune functionality.
Starting about a dozen years ago, many researchers started to believe that thymic involution could be reversed in older people and functioning of the adaptive immune system might thereby be restored.
This has been demonstrated to work in mice and dogs.
There are several possible known approaches for initiating such reversal.
It has been demonstrated in a clinical trial that administration of human growth hormone (hGH) restores thymic and T cell functionality in individuals infected with HIV. This possibility has been confirmed in other studies, but only in the context of HIV infection.,
Despite all this, approaches to halting or reversing age-related immune system decline in the translational medicine pipeline appear to be few and far between.
There are no new clinical trials relating to thymic involution or other approaches for halting or reversing age-related immune system decline on the books. In fact, there can’t be because aging is not officially a disease according to the FDA.
Most researchers now seem to have lost interest in thymus gland regeneration in healthy aging people – the literature focused directly on this topic was published in 2006 or earlier.
Some small biotech companies continue to work on the issue. Recombinant human IL-7 is being developed by Cytheris, Inc. right now for the treatment of thymic involution. It may be become FDA approved in the next 2-3 years. A small clinical feasibility trial using hGH may be in the works, according to a personal communication from Gregg Fahey, a long-time experimenter in this field.
New basic scientific discoveries potentially applicable to immune system regeneration continue to appear. There is new understanding of the molecular biology of thymic involution and thymic regeneration is known to be directly inducible via the FOXN1 transcription factor,
For individuals wishing to maintain their immune system functionality, there are a few things they can in fact do, some simple and practical, some more extreme. These are based on knowledge of many factors that can accelerate or slow down the age-related decline in functional immunity.
From a science viewpoint, the prospectus for partially or reversing age-related immune senescence is very good. It may be some time before practical and economic barriers that are stopping this from happening are overcome, however.
AGE-RELATED DECLINE IN THE INNATE IMMUNE SYSTEM
Athough immunosenescence is primarily a T cell problem and our central focus here is on the adaptive immune system, there is also related decline in the functionality of the innate immune system and many issues there that come with aging. The innate immune system includes neutrophils, monocytes/macrophages, microglia, dendritic cells, and natural killer cells. Here are the major aging-related impacts on the innate immune system:
A. Neutrophil changes- Neutrophils become less functional and undergo a lot of changes with aging. There may be a preserved neutrophil number or there can be a decrease in neutrophil number. Either way, they don’t work as well. There are major problems with decreased chemotaxis decreased opsonization, decreased phagocytosis, and decreased free radical production (peroxisomes). There is also a decrease in signaling molecule expression (GM-CSF-R, TLR-4, fMLP-R, TREM-1). There are also changes to the plasma membranes, including a reduced levels of cholesterol, increased membrane fluidity, dysregulation of receptor recruitment to lipid rafts, decreased receptor signaling (KPB, JAK, PI3K). There is a preserved expression of adhesion molecules however.
B. Monocyte/Macrophage changes- There is a clear INCREASE in the CD16+ monocyte/macrophage population with aging. This is a “pro-inflammatory phenotype”. There is also a decrease in antigen presentation and a decrease in phagocytic ability of the mononuclear cells. They also cannot make as many free radicals.
C. Dendritic Cell (DC) changes- Dendritic cells clearly decline in number with aging. They make less IL-12, which is an anti-inflammatory cytokine. There is also a decrease in the subpopulation of TLR-7 or TLR-9 cells.
D. Natural Killer cell (NK) changes- Overall increase in percentage and absolute NK cell number. There is an increase in some NK cell populations and a decrease in other NK cell populations, but overall, the absolute number increases with aging. However, there is a decrease in function of NK cells with aging. There is clearly a decrease in CD56+ subpopulation, which is an immunoregulatory subtype. However, there is an increase in the CD56- subpopulation, which are the cytotoxic NK cells. There is a preserved production of IFN-gamma. There is a decrease in chemokine production.
Age-related changes in some of these cell types can also impact on the adaptive immune system. For example, dendritic cells serve for communication between the innate and adaptive immune systems, and can activate lymphocytes and initiate other activities of the adaptive immune system.
AGE-RELATED DECLINE IN THE ADAPTIVE IMMUNE SYSTEM
The decline in functionality of the adaptive immune system is mainly dueto decline in numbers of naive T cells corresponding to increase in numbers of memory T cells.
Put very simply: relatively early in life the thymus makes what is supposed to be a lifelong supply of naïve T cells out of precursor cells in the bone marrow. When those naïve T cells encounter a foreign invader such as many bacteria, viruses or cancer cells, they convert to being memory cells which can recognize the same invader and speed up the defense against it later in life. The thymus essentially begins to shut down after adolescence (a process called thymic involution), meaning that naïve T cells can continue to divide but are no longer generated in the thymus. So as people age the population of naïve T cells become smaller and smaller and older and the population of memory cells becomes larger and larger reflecting the pathogens that have been encountered. More exactly:
“Memory T cells are a subset of infection- as well as potentially cancer-fighting T cells (also known as a T lymphocyte) that have previously encountered and responded to their cognate antigen; thus, the term antigen-experienced T cell is often applied. Such T cells can recognize foreign invaders, such as bacteria or viruses, as well as cancer cells. Memory T cells have become “experienced” by having encountered antigen during a prior infection, encounter with cancer, or previous vaccination. At a second encounter with the invader, memory T cells can reproduce to mount a faster and stronger immune response than the first time the immune system responded to the invader. This behaviour is utilized in T lymphocyte proliferation assays, which can reveal exposure to specific antigens(ref).”
“Thymic function decreases with age. (a) Top panel shows cytokeratin-immunostained and hematoxylin & eosin (H&E)-counterstained thymus sections from healthy young and aged human donors. In the aged thymus, the perivascular space (P) is increased in size and the thymopoietic cortex (C) and medulla (M) are significantly constricted. Lower panel shows flow cytometric analysis of CD4 and CD8 on thymocytes isolated from healthy young and aged human thymus donors; a clear reduction in double positive (DP) CD4+CD8+ T cells be seen in aged individuals. (b) sjTREC (single joint T-cell receptor excision circle) measurement per microgram of circulating peripheral blood mononuclear cell (PBMC) DNA from healthy human donors ranging from <1 to ∼80 years of age demonstrates a steady decline of naïve T-cell output with age. (c) Human thymus tissue from a 78-year-old female. Tissue was immunostained with anti-cytokeratin and H&E counterstained to show remaining islands of cortical and medullary tissue (blue). (d) Islands of cortical and medullary tissue in aged thymus show signs of active thymopoiesis by CD1a and Ki67 immunostain.”
Thymic involution: “One of the major characteristics of vertebrate immunology is thymic involution, the shrinking of the thymus with age, resulting in changes in the architecture of the thymus and a decrease in tissue mass.[1] This process is a conserved sequence or (orthologous sequences) in almost all vertebrates, from birds, teleosts, amphibians to reptiles, though the thymus of a few species of sharks are known not to involute.[1][2] T-cells are named for the thymus where T-lymphocytes migrate from the bone marrow to mature. Its regression has been linked to the reduction in immunosurveillance in the elderly.[3] Though thymic involution has been linked to senescence, it is not induced by senescence as the organ starts involuting from a young age [4] – as early as the first year of life in humans(ref).[5] ”
“The ability of the immune system to mount a strong protective response depends on the receptor diversity of naive T-cells (TCR). Thymic involution results in a decreased output of naïve T lymphocytes – mature T cells that are tolerant to self antigens, responsive to foreign antigens, but have not yet been stimulated by a foreign substance. In adults, naïve T-cells are hypothesized to be primarily maintained through homeostatic proliferation, or cell division of existing naïve T cells. Though homeostatic proliferation helps sustain TCR even with minimal to nearly absent thymic activity, it does not increase the receptor diversity.[11] For yet unknown reasons, TCR diversity drops drastically around age 65.[11] Loss of thymic function and TCR diversity is thought to contribute to weaker immunosurveillance of the elderly, including increasing instances of diseases such as cancers, autoimmunity, and opportunistic infections(ref).”
Quoting further from the earlier blog entry “One of the hallmark of advanced aging is weakening adaptive immune systems, often referred to as immunosenescence. “These age-associated immune dysfunctions are the consequence of declines in both the generation of new naïve T and B lymphocytes and the functional competence of memory populations(ref).” This decline appears to be due to a variety of causes. For one matter, thymic involution starts at an early age. “The thymus begins to shrink (atrophy) after adolescence. By middle age it is only about 15% of its maximum size(ref). There are strong age-related changes in hormone production(ref). Older people may take anti-inflammatory medications like prednisone which inhibit immune function. And changes in patterns of epigenetic markers alter gene activation so as to reduce responsiveness of T cells with age.”
“In particular, homeostatic mechanisms related to naïve T-cells tend to become deregulated with advancing age in primates and humans. A 2011 report Age-related deregulation of naive T cell homeostasis in elderly humansconcludes “Our results show that lower naive T cell numbers were associated with a lower thymic function and higher activation and proliferating naive T cell levels. We then analyzed sjTREC numbers and relative telomere length from sorted naive T cells. Our results show that the aberrant activation and proliferation status was related to lower sjTREC numbers (a peripheral proliferation marker) and both, higher CD57 expression levels and shortened telomeres (replicative senescence-related markers). Elderly individuals show a greater contraction of the CD8 naive T cell numbers and all homeostatic alterations were more severe in this compartment. In addition, we found that low functional thymus show a CD4-biased thymocyte production. Taken together, our results suggest a homeostatic deregulation, affecting mostly the naive CD8 T cell subset, leading to the accumulation of age-associated defects in, otherwise, phenotypically naive T cells.”
FACTORS RELATED TO FUNCTIONAL DECLINE IN ADAPTIVE IMMUNITY
These can be many, including problems with the haematopoietic precursor bone marrow cells, problems in the thymus, and existence of various triggers for thymic involution including sex hormones and certain diseases.
Impaired hematopoietic stem cells
A dramatic change in the immune system with aging is in the naive T cell compartment. Part of this is due to the inability to generate lymphoid progenitors by functionally impaired haematopoietic stem cells, the bone marrow cells that are processed by the thymus to make naïve T cells.. This impaired hematopoietic stem cell problem has been explained to be due to a deficiency in the capacity to repair DNA damage.
The 2007 publication Deficiencies in DNA damage repair limit the function of haematopoietic stem cells with age reports “A diminished capacity to maintain tissue homeostasis is a central physiological characteristic of ageing. As stem cells regulate tissue homeostasis, depletion of stem cell reserves and/or diminished stem cell function have been postulated to contribute to ageing. It has further been suggested that accumulated DNA damage could be a principal mechanism underlying age-dependent stem cell decline. We have tested these hypotheses by examining haematopoietic stem cell reserves and function with age in mice deficient in several genomic maintenance pathways including nucleotide excision repair, telomere maintenance and non-homologous end-joining. Here we show that although deficiencies in these pathways did not deplete stem cell reserves with age, stem cell functional capacity was severely affected under conditions of stress, leading to loss of reconstitution and proliferative potential, diminished self-renewal, increased apoptosis and, ultimately, functional exhaustion. Moreover, we provide evidence that endogenous DNA damage accumulates with age in wild-type stem cells. These data are consistent with DNA damage accrual being a physiological mechanism of stem cell ageing that may contribute to the diminished capacity of aged tissues to return to homeostasis after exposure to acute stress or injury.” Other publications from the same 2007-2008 era with similar themes include:
There are many more-recent articles relating to the functionality of aging and inadequate DNA repair such as can be found in this list. We comment that recent research suggests that a limiting factor in DNA repair in older people may be inadequate nuclear supplies of NAD+ — a situation that might be addressed in older people by supplementation with a NAD+ precursor such as nicotinamide riboside or nicotinamide mononucleotide. Because that subject is part of a larger picture of aging also involving the mitochondria, metabolism and ROS, we will deal with the intricacies of it in a separate blog entry that should be published shortly.
Thymic involution
This is a process that starts at age 1 and results in complete involution by 60 years of age for most people. As a result, 80% of the lymphocyte pool is depleted. This dramatically impairs the adaptive immune system’s capability to respond to new antigens. This decrease is seen in lymphoid organs and in peripheral blood. It is more pronounced in CD8+ than CD4+ cells. For CD4+ cells, the CD31 surface antigen correlates with naive status. The newly emigrated thymic CD4+ cells are CD4+CD31+. With aging, this population declines in a direct correlation with thymic involution.
Again from the earlier blog entry: ‘The thymus is also extremely susceptible to diseases. Malnutrition, nutritional imbalances and other triggers of thymic involution. See The thymus is a common target organ in infectious diseases(2006)and The thymus is a common target in malnutrition and infection (2007). The 2010 publication Nutritional imbalances and infections affect the thymus: consequences on T-cell-mediated immune responsesreports “The thymus gland, where T lymphocyte development occurs, is targeted in malnutrition secondary to protein energy deficiency. There is a severe thymic atrophy, resulting from massive thymocyte apoptosis (particularly affecting the immature CD4+CD8+ cell subset) and decrease in cell proliferation. The thymic microenvironment (the non-lymphoid compartment that drives intrathymic T-cell development) is also affected in malnutrition: morphological changes in thymic epithelial cells were found, together with a decrease of thymic hormone production, as well as an increase of intrathymic contents of extracellular proteins. Profound changes in the thymus can also be seen in deficiencies of vitamins and trace elements. Taking Zn deficiency as an example, there is a substantial thymic atrophy. Importantly, marginal Zn deficiency in AIDS subjects, children with diarrhoea and elderly persons, significantly impairs the host’s immunity, resulting in an increased risk of opportunistic infections and mortality; effects that are reversed by Zn supplementation. Thymic changes also occur in acute infectious diseases, including a severe thymic atrophy, mainly due to the depletion of CD4+CD8+ thymocytes, decrease in thymocyte proliferation, in parallel to densification of the epithelial network and increase in the extracellular matrix contents, with consequent disturbances in thymocyte migration and export. In conclusion, the thymus is targeted in several conditions of malnutrition as well as in acute infections. These changes are related to the impaired peripheral immune response seen in malnourished and infected individuals. Thus, strategies inducing thymus replenishment should be considered as adjuvant therapeutics to improve immunity in malnutrition and/or acute infectious diseases.”
Factors that accelerate the functional decline in Adaptive Immunity
CMV viral infections appear to be one the biggest factors that accelerates immune decline. Oxidative stress appears to be another major cause. The administration of sex steroid exogenous hormone replacement therapy also hastens immune aging. Smoking is another factor that accelerates immune senescence. “Among additional triggers for thymic involution are the presence of beta catenin, an oncogene(ref) and the presence of T-cell lymphoma(ref)(ref).”
Sex hormones and the thymus
It appears that sex hormones are implicated in thymic involution and that for males castration may be an effective anti-aging strategy, not that we recommend it. See for example the 2008 article The role of sex steroids and gonadectomy in the control of thymic involution. You can also check out articles in this list that relate hormones to thymic finctionality. Preservation of thymic activity may explain why Korean court eunuchs may have lived so long.. “They found that the average life span for a Korean court eunuch was about 70 years, plus or minus 1.76 years. That was 14.4 to 19.1 years longer than their average non-eunuch contemporaries, who tended to live between 50.9 and 55.6 years(ref)”. We note that this is a greater life span lengthening effect than any other intervention known in humans so far.
Decline in hGH and increases in sex steroids and glucocorticoids cause thymic involution => decline in Naive cells and increase in Memory cell senescence
Some authors have stated that involution of the mammalian thymus is one of the leading regulators of aging. In 1985, Steinman showed that thymic function actually starts to decline in the first year of life, but the dramatic involution of the thymus starts during puberty and is clearly driven by sex steroids. The block occurs at the TN1 to TN2 stage in the thymocyte differentiation of stromal cells within the thymus. These cells are identified by surface markers (CD3, CD4, and CD8). Development through these early stages is dependent on IL-7. However other important cytokines also stimulate thymopoiesis, such as keratinocyte growth factor (KGF), GM-CSF,
However, it appears that while blocking sex hormones may delay thymic involution, it cannot prevent it from eventually happening. The2005 publication Reassessing the role of growth hormone and sex steroids in thymic involution reports “The concomitant decline in growth hormone (GH) and increase in sex steroid production with age is thought to be responsible for thymic involution. If changes in the production of these hormones trigger or sustain thymic involution, that process should be accelerated in little mice, which have a genetic deficiency resulting in reduced production of thymopoietic GH, and delayed in the hypogonadal strain, which fails to produce thymocytotoxic sex steroids. The results indicated that thymic involution in both strains progressed in a manner similar to their normal littermates. That blocking sex steroid production did not delay thymic involution was surprising since castration reportedly increases thymus cellularity. Re-examination of that phenomenon revealed that, while gonadectomy results in increased thymus size, its effects are transient, and the thymus ultimately undergoes involution. Taken together, these data suggest that age-related changes in the endocrine system do not underlie thymic involution.”
“The strength of the immune response declines with age: Multiple age-related changes can affect the composition and function of lymphocytes in secondary lymphoid tissues. CD4+ Th cells exhibit activation defects and increased differentiation into Th17 cells. CD8+ T cells undergo an oligoclonal expansion and loss of CD28 in humans and exhibit impaired function. The number of B cells that respond to influenza is reduced, and antibody avidity in response to carbohydrate antigens is diminished. In addition, the tissue environment includes an increased concentration of inflammatory cytokines, which may be produced by stromal elements, dendritic cells, or aging B and T cells. The increased number of memory cells that occupy tissue niches and the inflammatory milieu in turn may compromise the ability of naive B and T cells migrating from the bone marrow and thymus to lodge in the tissue. Together, these changes result in diminished immune function in the elderly. “
“Effects of aging on HSCs and lymphocyte progenitors: Lymphopoiesis in the young (left) is characterized by robust B and T cell production in the bone marrow and thymus. The pool of HSCs includes a relatively high number of lymphoid-biased stem cells that efficiently generate lymphoid progenitors with high proliferative potential. However, with increasing age (right), the number of lymphoid-biased HSCs declines and myeloid-biased stem cells predominate, contributing to the reduced numbers of lymphoid progenitors. In addition, B cell progenitors in the bone marrow and T cell progenitors in the thymus exhibit reduced rates of proliferation and higher levels of apoptosis compared with their young counterparts. The increased expression of Ink4a and Arf in pro-B cells and Ink4a in ETPs contribute to this decreased proliferation/increased apoptosis. The decline in primary lymphopoiesis in turn results in a reduced number of naive cells that migrate to secondary lymphoid tissues such as the spleen. “
Reversing thymic involution as a strategy for restoring age-related decline in immune function
There appears to be wide agreement that thymic involution can be halted or reversed, This topic was treated in the earlier blog entry and again is the focus here. This diagram is also from the 2009 publication Thymic involution and immune reconstitution.
In part, the legend states “Treatment with thymostimulatory cytokines, such as keratinocyte growth factor (KGF), interleukin 7 (IL-7), growth hormone (GH), leptin and ghrelin, or sex steroid ablation therapy (SSA), can promote regeneration of an atrophic thymus—increasing total cellularity, restoring thymic architecture, increasing output of naïve T cells and rejuvenating the diversity of the peripheral TCR repertoire. This might occur by several mechanisms including an increase in thymus seeding by early T-lineage progenitors (ETPs) or increased proliferation and/or differentiation of triple negative (TN), double positive (DP) or single positive (SP) thymocytes.” That document also reports “Chronic thymus involution associated with aging results in less efficient T-cell development and decreased emigration of naïve T cells to the periphery. Thymic decline in the aged is linked to increased morbidity and mortality in a wide range of clinical settings. Negative consequences of these effects on global health make it of paramount importance to understand the mechanisms driving thymic involution and homeostatic processes across the lifespan. There is growing evidence that thymus tissue is plastic and that the involution process might be therapeutically halted or reversed. We present here progress on the exploitation of thymosuppressive and thymostimulatory pathways using factors such as keratinocyte growth factor, interleukin 7 or sex steroid ablation for therapeutic thymus restoration and peripheral immune reconstitution in adults.”
Strategies for slowing, halting or reversing thymic involution
A substantial body of research literature has emerged suggesting strategies for slowing, halting or reversing thymic involution. See the articles in this list. Some of these approaches were reviewed in the earlier blog entry. Here, we will focus mainly on more recent research, such as related to administration of the transcription factor Foxn1, and on use of human growth factor hGH..
“Selected strategies to rejuvenate the involuted thymus. The potential of several hormones and growth factors to rejuvenate the involuted thymus has been tested in various preclinical and clinical trials. Many of these factors can be grouped into three categories. Those in the first (i), such as IL-7, bind to progenitors in the bone marrow and thymus and have only modest effects on thymopoiesis. There is little evidence that IL-7 has effects on thymic stromal cells. Instead, the benefit of IL-7 may lie in its ability to stimulate peripheral T cell survival/expansion. The second category (ii) includes hormones such as GH that have been demonstrated in preclinical and clinical trials to stimulate thymopoiesis and increase thymic size. Many GH effects are mediated through induction of IGF-1. IGF-1 can bind to receptors on thymic stroma and thymocytes, although its actions are primarily mediated through effects on the former cells. Stromal cell–derived factors presumably then act on thymocytes (curved arrow). A third category of factors (iii), typified by FGF7, bind to stromal cells but not thymocytes. Stromal cell–induced factors then act on thymocytes (curved arrow), and we have recently demonstrated that effects include downregulation of Ink4a in ETPs (82). The thymopoietic effects of several additional factors have been evaluated, and recent detailed reviews should be consulted for more information (99, 100).”
The key role of FOXN1
Recent mouse-level studies indicate that a single transcription factor FOXN1 can both halt thymic involution and initiate regeneration of an involuted thymus. The FOXN1 gene exists in mice and humans and FOXN1 knockout mice have been used to study cancer and skin issues. More recently, a number of studies have appeared that relate FOXN1 to thymus gland development and regeneration(ref)(ref)(ref)
The 2011 publication relates to aged miceOverexpression of Foxn1 attenuates age-associated thymic involution and prevents the expansion of peripheral CD4 memory T cells. “The forkhead box n1 (Foxn1) transcription factor is essential for thymic organogenesis during embryonic development; however, a functional role of Foxn1 in the postnatal thymus is less well understood. We developed Foxn1 transgenic mice (Foxn1Tg), in which overexpression of Foxn1 is driven by the human keratin-14 promoter. Expression of the Foxn1 transgene increased the endogenous Foxn1 levels. In aged mice, overexpression of Foxn1 in the thymus attenuated the decline in thymocyte numbers, prevented the decline in frequency of early thymic progenitors, and generated a higher number of signal joint TCR excised circle. Histologic studies revealed that structural alterations associated with thymic involution were diminished in aged Foxn1 Tg. Total numbers of EpCAM+ MHC II+ and MHC II(hi) thymic epithelial cells were higher in young and old Foxn1Tg and more EpCAM+ MHC II(hi) TEC expressed Ki-67 in aged Foxn1Tg compared with WT. Furthermore, Foxn1Tg displayed a significant reduction in the expansion of splenic CD4+ memory compartments and attenuated the decline in CD4+ and CD8+ naive compartments. Our data indicate that manipulation of Foxn1 expression in the thymus ameliorates thymopoiesis in aged mice and offer a strategy to combat the age-associated decline in naive T-cell production and CD4 naive/memory ratios in the elderly.
The 2013 publication Enhancing T lineage production in aged mice: a novel function of Foxn1 in the bone marrow nichereports “Foxn1 is essential for thymic organogenesis and T lymphopoiesis. Whereas reduced Foxn1 expression results in a decline in T lymphopoiesis, overexpression of Foxn1 in the thymus of a transgenic mouse model (Foxn1Tg) attenuates the age-associated decline in T lymphopoiesis. T lymphopoiesis begins with early T cell progenitors (ETP), derived from multipotent progenitors (MPP) in the bone marrow (BM). A decline in MPP and ETP numbers with age is thought to contribute to reduced T lymphopoiesis. Previously, we showed that reduced ETP number with age is attenuated in Foxn1 transgenic (Tg); whether the effect is initiated in the BM with MPP is not known. In this study, we report that Foxn1 is expressed in wild-type BM and overexpressed in Foxn1Tg. With age, the number of MPP in Foxn1Tg was not reduced, and Foxn1Tg also have a larger pool of hematopoietic stem cells. Furthermore, the Foxn1Tg BM is more efficient in generating MPP. In contrast to MPP, common lymphoid progenitors and B lineage cell numbers were significantly lower in both young and aged Foxn1Tg compared with wild type. We identified a novel population of lineage(neg/low), CD45(pos) EpCAM(pos), SCA1(pos), CD117(neg), CD138(neg), MHCII(neg) cells as Foxn1-expressing BM cells that also express Delta-like 4. Thus, Foxn1 affects both T lymphopoiesis and hematopoiesis, and the Foxn1 BM niche may function in skewing MPP development toward T lineage progenitors.
Finally, the 2014 publicationRegeneration of the aged thymus by a single transcription factor reports: “Thymic involution is central to the decline in immune system function that occurs with age. By regenerating the thymus, it may therefore be possible to improve the ability of the aged immune system to respond to novel antigens. Recently, diminished expression of the thymic epithelial cell (TEC)-specific transcription factor Forkhead box N1 (FOXN1) has been implicated as a component of the mechanism regulating age-related involution. The effects of upregulating FOXN1 function in the aged thymus are, however, unknown. Here, we show that forced, TEC-specific upregulation of FOXN1 in the fully involuted thymus of aged mice results in robust thymus regeneration characterized by increased thymopoiesis and increased naive T cell output. We demonstrate that the regenerated organ closely resembles the juvenile thymus in terms of architecture and gene expression profile, and further show that this FOXN1-mediated regeneration stems from an enlarged TEC compartment, rebuilt from progenitor TECs. Collectively, our data establish that upregulation of a single transcription factor can substantially reverse age-related thymic involution, identifying FOXN1 as a specific target for improving thymus function and, thus, immune competence in patients. More widely, they demonstrate that organ regeneration in an aged mammal can be directed by manipulation of a single transcription factor, providing a provocative paradigm that may be of broad impact for regenerative biology.”
There are many issues connected with non-thymic actions of FOXN1, and questions as to whether the mouse finding can be applied to thymus regeneration in humans. The 2012 publication Insights on Foxn1 Biological Significance and Usages of the “Nude” Mouse in Studies of T-Lymphopoiesisrelates: “FoxN1 is a transcription factor whose functions are executed by targeting other genes through its DNA binding domain. Therefore, to understand its functional mechanisms in determining its target genes is important. However, the precise target genes that are regulated by FoxN1 remain ill defined, mostly due to technical difficulties in precisely isolating enough physiologically intact TECs at certain developmental stages. — Recent progress using advanced technology to study FoxN1‘s roles in the thymus shows that FoxN1 regulates not only TEC patterning in the fetal stage but also TEC homeostasis in the postnatal thymus. Comparing the thymus with the skin, FoxN1 has its own distinct roles and impacts on organs in the generation and maintenance of three-dimensional microstructure and pigmentation, respectively. FoxN1‘s role in the neuron has been brought up, but is still obscure. There is still plenty of room to apply nude and secondary nude (conditional FoxN1 gene knockout) mouse models in studies of immunology, hematology, and tumorgenesis. The functional mechanisms of FoxN1‘s collaborative roles with other genes during thymic development and aging remain to be further determined.”
Other publications related to FOXN1 and the thymus are
My assessment is that it will be some time before a FOXN1 therapy for reversing human thymic involution reaches the clinical trial stage, if ever. We are more likely to see a clinical trial relating to FOXN1 as a therapy for averting immunosenescence in HIV patients. Reversing thymic involution would still be the trial target, but the context for testing would be limited to HIV-infected people.
Human growth hormone treatment for reversing thymic involution
The use of growth hormone to restore thymic functionality in animal models goes back to 1986, when the implantation of pituitary adenoma cells secreting GH into rats was seen to induce thymic growth. See the publication GH3 pituitary adenoma cells can reverse thymic aging in rats. The following year, growth hormone treatment in dogs was shown to increase thymulin production. See Growth hormone treatment stimulates thymulin production in aged dogs. In 2003 Gregg Fahey reported partial thymic regeneration due to hGH administration in a single human being (thought to be himself)(ref). .
IGF-1 is induced by hGH
The hormone that actually induces thymulin is IGF-1, which is produced by the liver in response to hGH release from the pituitary. Such hGH release is thought to be the major reason why the hormone thymulin declines with aging. GH induces both bone marrow rejuvenation and thymic rejuvenation. This effect on the bone marrow is normally mediated by CSF-1 and IL-3, which both induced expression and synthesis of IGF-1 locally within the bone marrow by bone marrow cells. However, adding exogenous IGF-1 to bone marrow cells decreases apoptotic cell death by 50%. See the 1996 document Growth Hormone, Growth Factors and Hematopoiesis.
Safety of hGH or IGF-1 treatment
hGH administration is more often thought to shorten rather than lengthen lilespans(ref)(ref)(ref).
The reader may recall that in many of our blog entries we have discussed the Dr. Jekyll and Mr, Hyde nature of IGF-1. On the one hand it is essential for growth and wound healing. On the other hand, IGF-1 expression promotes many disease processes and hallmarks of aging. There seems to be a lot of agreement that to slow or prevent aging, it is desirable to minimize IGF-1 expression and therefore not induce it with hGH. In fact, you want to suppress expression of IGF-1. See the blog entry Longevity and the GH–IGF Axis. Therefore it might seem that using hGH for thymic regeneration may involve both life extending and life-shortening aspects. It could be, however, that hGH induces a biphasic response curve and that the low dose required for thymic regeneration is safe and beneficial.
Clinical trials of hGH for reversing thymic involution
A highly relevant clinical trual Growth Hormone to Increase Immune Function in People With HIV was initiated in 2003 by the National Institutes of Allergy and Infectuous Diseases. As reported in the 2008 publication Growth hormone enhances thymic function in HIV-1-infected adults: “Growth hormone (GH) is an underappreciated but important regulator of T cell development that can reverse age-related declines in thymopoiesis in rodents. Here, we report findings of a prospective randomized study examining the effects of GH on the immune system of HIV-1-infected adults. GH treatment was associated with increased thymic mass. In addition, GH treatment enhanced thymic output, as measured by both the frequency of T cell receptor rearrangement excision circles in circulating T cells and the numbers of circulating naive and total CD4(+) T cells. These findings provide compelling evidence that GH induces de novo T cell production and may, accordingly, facilitate CD4(+) T cell recovery in HIV-1-infected adults. Further, these randomized, prospective data have shown that thymic involution can be pharmacologically reversed in humans, suggesting that immune-based therapies could be used to enhance thymopoiesis in immunodeficient individuals.”
“Comparison of changes in the GH arm versus the observational control arm over the first year of the study showed that GH treatment was associated with significant increases in the percentage (A) and absolute count (B) of CD4+ T cells (red). (C) Comprehensive regression analysis, including crossover data of GH treatment in observational controls, showed that GH treatment (circles) was associated with significant increases in CD4+ T cell percentage and the CD4+/CD8+ T cell ratio when compared with no GH. Increases in the absolute count of CD4+ T cells trended toward statistical significance in this analysis. There were no remarkable GH-associated changes in the percentage or absolute count of CD8+ T cells. Estimated changes with 95% CIs are shown. Regression analysis estimated the effects of 1 year of GH treatment compared with changes over 1 year in the absence of GH. Median values are shown in A and B. CIs and additional data are shown in Tables Tables22 and and3.3. *P < 0.05 for comparison of GH versus no GH.”
A subsequent 2013 publication Low-dose growth hormone for 40 weeks induces HIV-1-specific T cell responses in patients on effective combination anti-retroviral therapyreports: “Recombinant human growth hormone (rhGH) administered to combination anti-retroviral therapy (cART)-treated human immunodeficiency virus-1 (HIV-1)-infected individuals has been found to reverse thymic involution, increase total and naive CD4 T cell counts and reduce the expression of activation and apoptosis markers. — Here we report that administration of low-dose rhGH over 40 weeks with effective cART resulted in greater improvement of T lymphocyte function than observed with cART alone, and provide further evidence that such an approach could also reduce levels of immune activation. “
Do we think that the positive benefits seen in HIV-infected people would also show up in older people who have age-related thymic involution?
In short, yes. Why not?
So,research on reversal of decline in thymus and lymphocyte function via hGH administration seems to be alive and well in the subdomain of treatment of patients infected with HIV. Not so in the case of aging. The initial mouse literature on use of GH for reversal of thymic involution go back to 2002 and literature discussions of use on HGH as an effective human immune-extending therapy seem to have petered off around 2006. There seems to be no more-recent research literature related directly to hHG administration for reversing thymic involution in normal aging individuals – although this idea continues to be discussed in some anti-aging blogs.
Based on private communications, I think there may be a small feasibility clinical trial related to hGH restoration of aging involuted thymic glands proposed by a biotech company in the near future. Other than for that, I do not forsee much real action here. The issue is that FDA-authorized clinical trials can only be for disease conditions, and aging is not considered to be a disease.
The rest of this document is concerned with additional factors that might contribute to thymic involution or reconstitution, and with some possible practical applications of these.
Ghrelin - In 1999, an endogenous ligand for the previously discovered “growth hormone secretagogue receptor” (GHS-R) was discovered. This molecule is now called Ghrelin and activates the GHS-R, which is a specific G protein-coupled receptor. Ghrelin is commonly called “the hunger molecule.” See the blog entry Ghrelin, hunger, obesity and aging. There is both an acylated ghrelin and a des-acyl form of gherkin circulating in the plasma. The des-acyl ghreline does not bind to GHS-R1a, but both the acylated and des-acyl form binds to common sites on cardiomyocytes and endothelial cells. They work similarly to IGF-1 to prevent apoptosis in cells. Ghrelin is able to strongly stimulate the release of hGH from the pituitary by binding to the GHS-R. Ghrelin is produced by the stomach, but it is also produced elsewhere in the body. Ghrelin inhibits leptin and induces hunger. Ghrelin is actually produced by human T cells, monocytes, and dendritic cells. Ghrelin has potent inhibitory effects on pro-inflammatory cytokine production by activated immune cells, such as T cells, monocytes, and dendritic cells. Ghrelin significantly increases the cellularity of the thymus in older mice. Ghrelin also increased the engraftment of a bone graft. Ghrelin looks like a promising new approach to stimulating the thymus.
2. Gonadectomy – We have already mentioned castration and the affect of sex hormones on thymic vitality Over 100 years ago, gonadectomy in both male and female animals was shown to delay, but not prevent thymic involution. When done later in life, gonad removal induces a profound thymic rejuvenation. This has been done experimentally in animals and has been observed clinically in humans who underwent orchiectomy or oophorectomy for cancer. The molecular mechanisms of how castration regenerates the thymus are initiated in the “immature triple-negative compartment (CD3-, CD4-, CD8-) and early T lineage progenitors (ETP = Lin-, CD127-, CD25-. CD44+, CD117+). With aging, there is a reduction in the number of ETPs in the CD127- TN1 compartment. Castration restored the number of triple negative cells and the ETPs. The effects of castration in male mice are rapid. Within 6 hours, the levels of testosterone drop to 1% of normal levels. Within 10 days, there is a regeneration of the thymus, producing a cellularity that resembles a 2 month old mouse.
Pardoxically, gonadectomy temporarily increases thymic size, but then with time the thymus involutes again with aging. This is due to the decline in hGH that occurs with aging. Thus unless both the effects of sex steroids and the age-related decline in hGH are addressed, there will be a only a transient benefit to either gonadectomy or hGH administration in old age.
Chemical gonad hormonal ablation. – Chemical ablation of sex steroids also induces a profound thymic rejuvenation. This is seen with the use of the drug that interferes with luteinizing hormone releasing hormone (LHRH, aka GnRH). Both LHRH antagonists and GnRH antagonists have been used to induce a chemical gonad hormonal ablation. Initially, only a GnRH agonist was available for prostate cancer (Luprolide, aka Lupron), but recently GnRH antagonists have also been developed. Paradoxically, however, GnRH agonists reduce IGF-1. This is a surprising finding, since IGF-1 actually induces thymic regrowth. Thus GnRH agonists may not help with thymic rejuvenation. (ref)(ref)(ref)(ref)(ref)
Pregnancy - Pregnancy is associated with high levels of estrogen and progesterone. Accompanying these high levels of sex hormones, there is a dramatic thymic involution that occurs in the mother. In mouse models, the amount of thymic involution varies from 45% to 80%. Estrogen blocks early T cell development in the thymus. Estrogen induces a dramatic reduction in the thymic size and cellularity. All subsets of T cells decline (CD4 and CD8 cells). The triple negative cells (CD3-, CD4-, CD8-) did not progress from TN1 to TN2 developmental phases. Progesterone does not block T cell progression in the thymus, however. (ref)(ref)(ref)
Adrenal corticosteroids – Suprarenalectomy (adrenalectomy) in rabbits was done 90 years ago and showed a dramatic effect in arresting thymic involution and results in dramatic thymic regeneration. Involution is permanently delayed, but in the rabbits there was regrowth of accessory adrenal gland tissue. Unfortunately, in the adrenalectomized rabbits, there was a very high mortality rate if future surgery on the rabbits was performed. This is thought to be due to the lack of stress hormones needed to cope with these major stressors, such as surgery. However, if gonadectomy and adrenalectomy were performed, permanent prevention of thymic involution occurred.
Chronic glucocorticoid administration has also been shown to induce thymic involution. This occurs because glucocorticoids induce apoptosis in double positive thymic T cells (CD4-, CD8- cells). The mechanism for this is that glucocorticoids like cortisol migrate into the cytoplasm of the cell where it binds to the glucocorticoid receptor (GR). This induces a nuclear translocation of the cortisol/GR complex where it binds to hormone response elements (HREs) in promoter regions of genes. This direct ligand/GR binding to HREs accounts for the positive control of gene expression (i.e. turning genes on) but does not account for the negative transcriptional control (turning genes off). Negative transcriptional control of gene expression by ligand/GR is mediated by interference with the cAMP-responsive transcription factor, CREB, and interference with transcription factor AP-1. In the case of NF-kB, cortisol/GR “turns off” NF-kb by inducing IkBalpha, a specific NF-kB inhibitor.(ref)
6. Zinc deficiency – Zinc deficiency plays a major role in thymic involution later in life (not puberty). Two distinct epithelial cell populations in the thymus produce a hormone called thymulin (aka Thymic Factor). It is a nonapeptide and requires Zinc for its biological activity. Thymulin is important in T-cell differentiation and with the enhancement of T cell and NK cell action. The lack of Zinc at the thymulin receptors on T-cells is a major cause of thymic involution. In studies done in mice, oral zinc supplementation in old mice for 1 month induced a complete recovery of crude zinc balance from negative to positive values. A full recovery of thymic function with regrowth of the thymus occurred with partial restoration of immune efficiency, as measured by NK activity. The 1995 publication Reversibility of the thymic involution and of age-related peripheral immune dysfunctions by zinc supplementation in old micereported “A full recovery of thymic functions with a regrowth of the organ and a partial restoration of the peripheral immune efficiency, as measured by mitogen responsiveness (PHA and ConA) and natural killer cell (NK) activity, were observed after zinc supplementation. These findings clearly pin-point the relevance of zinc for immune efficiency and suggest that the age-related thymic involution and peripheral immunological dysfunctions are not intrinsic and irreversible events but are largely dependent on the altered zinc pool.”(ref)
Magnesium deficiency - Mg++ deficiency has also been shown to accelerate thymic involution. This appears to be due to enhanced apoptosis and thymocyte sensitivity to oxidative stress. Under experimental conditions where magnesium deficiency is induced in rats, thymic involution started to occur within a few weeks after the establishment of magnesium deficiency. Up until recently, the molecular mechanism by which magnesium deficiency induced thymic involution was not clear. Today there is evidence that this may be mediated by inflammation, which induces an increase in IL-6. An increase in IL-6 occurs within 4 days following the establishment of Mg deficiency.(ref)(ref)(ref)
Clinical Significance of the Above
Q: Why does hormone replacement therapy with estrogen or testosterone in middle age cause thymic collapse?
A: When testosterone or estrogen therapy is given to men and women after age 50, there is an almost complete collapse of whatever thymus is left after the 30-40 years of thymic involution that has occurred since puberty. This is due to apoptosis of the thymic cortical thymocytes, primarily the immature, pristine clonal stem cells that produce the upstream triple negative thymocytes. Thymic expression of androgen receptors increases with age. This may be why the thymus involutes without any exogenous sex hormone replacement therapy. It is unclear if the thymic involution is due only to apoptosis of thymocytes or if it is also due to the apoptosis of thymic stromal cells. Most believe that it is both.(ref)
Q What might be done about thymic involution?
A: The simplest option would be to make sure that there is no deficiency in Zinc or Magnesium, since Zn++ is required for thymulin binding to the thymulin receptors. Magnesium should reduce IL-6 mediated thymic involution. Another more drastic option would be to undergo castration or oophorectomy, but this will only have a transient effect and may not be desirable for many obvious reasons!
Another option would be to administer Keratinocyte Growth Factor (KGF). KGF is already FDA-approved for oral mucositis prophylaxis in patients receiving myeloablative therapies before hematopoietic stem cell transplantation.
Another option would be to administer IL-7. IL-7 augments the survival of triple negative thymocytes and single positive thymocytes. Administration of IL-7 after periods of immunodeficiency enhances T-cell reconstitution. This occurs via increased thymic T cell development.
Another option discussed above would be to administer hGH. This has been shown to effectively regenerate the thymus in AIDS patients. However even hGH administration would not last if the issue of sex hormones was not addressed. For this reason another option would be to do chemical down-regulation of sex hormones, such as the use of Luprolide. Unfortunately, luprolide decreases IGF-1, so this may have an antagonistic effect on the treatment of thymic involution with hGH.
Final questions and answers
Getting back to the issue posed by the title of this blog,
Q Do I think it is feasible to develop an effective therapy for reversing age-related immune senescence, at least partially if not fully?
Yes, with fairly high confidence. The required knowledge has been around for some time and the clinical trial for HIV patients establishes feasibility. The knowledge has simply not moved down the pipeline to where it is applied.
Q So, what has been stopping this from happening?
One major barrier is that hGH and IL-7 are natural substances that can’t be patented. So, little money is to be made selling them and there is no incentive for a pharma or biotech company to get involved in a big way. Not so for recombinant versions, but these may be little better than the natural ones. Another barrier common to all therapies aimed squarely at aging is that you can’t conduct a FDA-approved clinical trial for a purely aging indicator. Finally, in the case of hGH, there is the possible issue of adjusting dosage so as to minimize possible pro-disease and pro-aging side effects.
Then, will it happen?
Yes, I think so as the market for scientifically-validated anti-aging interventions gathers steam.
MEDICAL DISCLAIMER
FROM TIME TO TIME, THIS BLOG DISCUSSES DISEASE PROCESSES. THE INTENTION OF THOSE DISCUSSIONS IS TO CONVEY CURRENT RESEARCH FINDINGS AND OPINIONS, NOT TO GIVE MEDICAL ADVICE. THE INFORMATION IN POSTS IN THIS BLOG IS NOT A SUBSTITUTE FOR A LICENSED PHYSICIAN’S MEDICAL ADVICE. IF ANY ADVICE, OPINIONS, OR INSTRUCTIONS HEREIN CONFLICT WITH THAT OF A TREATING LICENSED PHYSICIAN, DEFER TO THE OPINION OF THE PHYSICIAN. THIS INFORMATION IS INTENDED FOR PEOPLE IN GOOD HEALTH. IT IS THE READER’S RESPONSIBILITY TO KNOW HIS OR HER MEDICAL HISTORY AND ENSURE THAT ACTIONS OR SUPPLEMENTS HE OR SHE TAKES DO NOT CREATE AN ADVERSE REACTION.
 Image may be NSFW.
Image may be NSFW. Image may be NSFW.
Image may be NSFW.